| Compléments sur les récepteurs couplés aux protéines G (RCPG) |
| Tweet |
|
|
1. Généralités sur les récepteurs couplés aux protéines G 2. Les principales classes (ou familles) de récepteurs couplés aux protéines G 3. Classification et des RCPG via leur spécificité de fixation de ligand 4. Détails de la structure conservée des récepteurs couplés aux protéines G |
5. Comparaison des conformations active et inactive d'un RCPG modèle 6. Comparaison des RCPG de la classe A et de la classe B 7. Liens Internet et références bibliographiques |
|
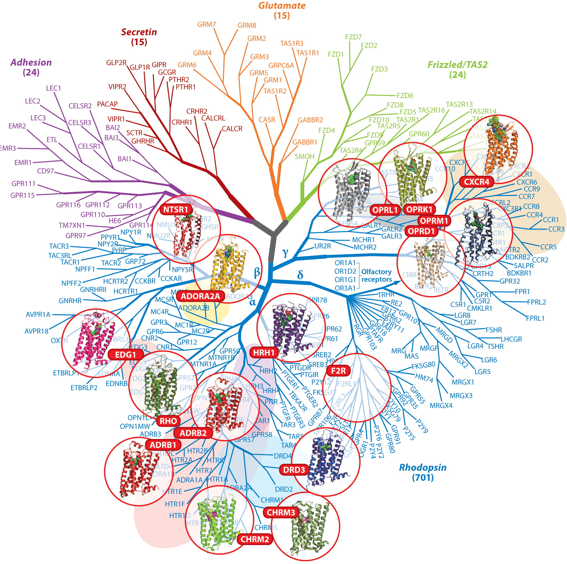
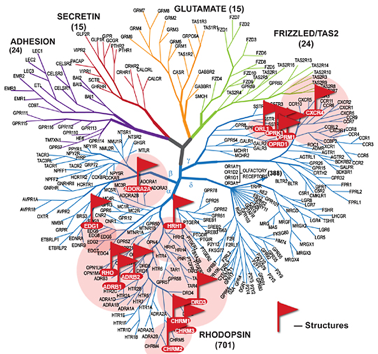
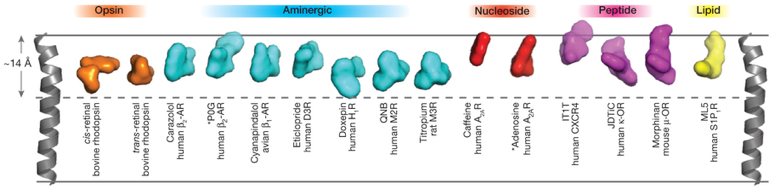
1. Généralités sur les récepteurs couplés aux protéines G On recense plus de 1000 récepteurs couplés aux protéines G (RCPG) (voir un cours). Les séquences codantes de leur gènes représentent plus de 1% du génome : plus de 800 gènes ont été identifiés chez l'homme (voir Fredriksson et al., 2003). Quelques représentants des différentes classes de RCPG dont la structure a été déterminée :
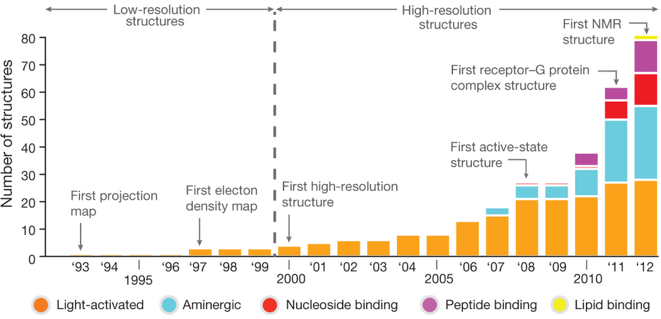
Source : Katritch et al. (2013) Figure ci-dessous : évolution du nombre et de la qualité des structures de GPCR déterminées par différentes techniques. Source : Venkatakrishnan et al. (2013) |
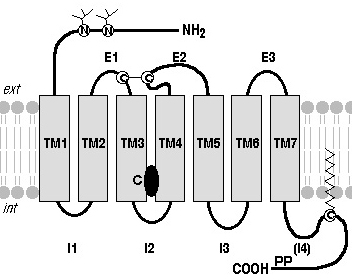
La structure globale de la grande majorité des RCPG est caractérisée par : Source : "Récepteurs couplés aux protéines G"
|
|
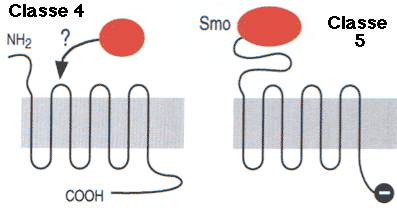
2. Les principales classes (ou familles) de récepteurs couplés aux protéines G Conformément aux directives de l'"International Union of Basic and Clinical Pharmacology", les RCPG non-sensoriels (c'est-à-dire à l'exclusion donc des récepteurs de lumière, odeur et goût) peuvent être classés, selon leurs propriétés pharmacologiques, en quatre grandes familles ou classes (selon les nomenclatures) :
Les GPCR chez l'homme sont activés à partir de différents sites de liaison de ligands endogènes dans le domaine transmembranaire (classe A) ou dans le domaine extracellulaire (classe C) ou dans les deux (classes B et F). Les classes ont une très faible similarité de séquence : une moyenne de 23 % pour les paires de classes croisées. |
| Classe de RCPG | Famille de RCPG | Exemple de ligands |
Classe A ("Rhodopsin like receptor family"). Ce sont de loin les plus nombreux : à ce jour plus de 660 gènes identifiés chez l'homme. |
||
Classe A / sous-type 1 |
Famille 1 / sous-groupe a | Rétinal, odorant, opiacés, β2-adrénergiques, dopaminergiques, histaminiques |
Classe A / sous-type 2 |
Famille 1 / sous-groupe b | bradykinine, angiotensine II, cytokines, thrombine |
Classe A / sous-type 3 |
Famille 1 / sous-groupe c | Hormones, glycoprotéiques |
| Classe B ("Secretin like receptor family") | Famille 2 | Calcitonine, sécrétine |
| Classe C ("Metabotropic glutamate/pheromone receptor family") | Famille 3 | Glutamate, acide γ-aminobutyrique (GABA), calcium, phéromones |
| Familles 4 et 5 | Phéromones / développement embryonnaire | |
| Voir la base de données dédiée aux RCPG : GPCRDB. | ||
|
Source : S. Martin Source : P. Sarret |
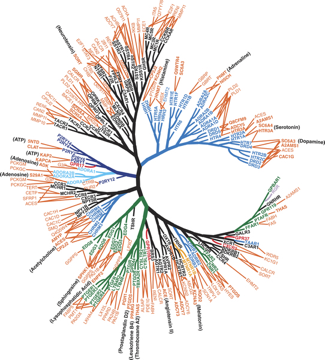
3. Classification des RCPG via leur spécificité de fixation de ligand Il existe de nombreux exemples de protéines ayant des similitudes structurales qui fixent des ligands différents et qui ont donc des fonctions différentes. A l'inverse, il existe de nombreux exemples de protéines ayant des similitudes fonctionnelles avec des structures différentes. Le nombre croissant de RCPG de différentes classes identifiés et de leur(s) ligand(s) permet, non pas de comparer les séquences protéiques, mais l'ensemble des ligands connus pour se fixer à un type de RCPG. La classification des protéines via leur similitude de liaison à un ou des ligand(s) est une approche alternative aux méthodes évolutives pour mieux comprendre la fonction des protéines et la transduction des signaux. La comparaison du dendrogramme basé sur la similarité des ligands à celui basé sur la similarité des séquences de RCPG de la classe A (figure de gauche ci-dessous), a permis d'identifier des RCPG éloignés du point de vue de leurs séquences mais qui ont des similitudes de spécificité de ligand. Ce genre d'approche a des applications dans la recherche de nouvelles cibles thérapeutiques et la mise au point de nouveaux médicaments.
Source : Lin et al. (2013) et GPCR network |
4. Détails de la structure conservée des récepteurs couplés aux protéines G La structure d'un RCPG peut être divisée en 3 régions :
a. Région extracellulaire et poche de fixation du ligand Les analyses des séquences montre une grande diversité dans les longueurs et les compositions en acides aminés de l'extrémité N-terminale et des boucles extracellulaires. Les structures de RCPG de la classe A révèlent 2 types de région extracellulaire : celles dont la poche de fixation du ligand est accessible à l'eau de ligands poche et celles dont la poche de fixation du ligand est inaccessible à l'eau (exemple : rhodopsine et récepteur S1P1). Figure ci-dessous : comparaison de la profondeur des poches de fixation des ligands des RCPG de la classe A. L'hélice transmembranaire TM4 est utilisée comme référence. Source : Venkatakrishnan et al. (2013) La profondeur de pénétration du ligand est la plus grande pour la doxépine pour le récepteur de l'histamine H1 et la moins grande pour la caféine pour le récepteur A2AR. Les étiquettes au dessus des structures indiquent la sous-classe des RCPG de la classe A. Une autre caractéristique de la région extracellulaire est la présence de ponts disulfures qui contribuent à la stabilité du récepteur. Le pont disulfure établi entre une Cys de TM3 (Cys3.25 - numérotation Ballesteros - Weinstein) et ECL2 semble hautement conservé dans la plupart des structures des RCPG (sauf S1P1). Ce pont disulfure TM3 - ECL2 ancre la région extracellulaire de l'hélice près du site de fixation du ligand et limite l'ampleur des changements conformationnels de cette région au cours de l'activation du récepteur. Plusieurs ECL3 contiennent un pont disulfure intra-boucle supplémentaire (motif CxnC) qui semble avoir une influence sur la fonction du récepteur en limitant les mouvements conformationnels de cette boucle. Par exemple, on a mis en évidence qu'une mutation faux-sens (C271R) de ce pont disulfure du récepteur de la mélanocortine-4 induit un dysfonctionnement du récepteur lié à l'obésité. Voir un article très détaillé sur la structure et les rôles des boucles extra-cellulaires : Wheatley et al. (2012) "Lifting the lid on GPCRs: the role of extracellular loops" Br. J. Pharmacol. 165, 1688 - 1703 |
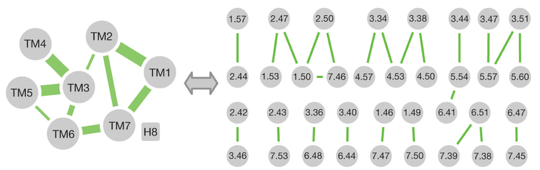
b. Repliement conservé de la région TM Le faisceau d'hélices TM sert à la communication entre la poche de liaison du ligand et la région de couplage aux protéines G. Bien que l'ensemble des RCPG partagent une architecture similaire de 7 hélices TM maintenue par des contacts au niveau de la structure tertiaire, leurs séquences sont diverses. Une analyse systématique des différentes structures de RCPG (incluant des RCPG dans l'état actif et inactif) a révélé un réseau consensus de 24 contacts inter-TM médiés par 36 acides aminés équivalents sur le plan topologique. L'importance de ces positions est confortée par le fait que des mutations dans 14 des 36 positions entraînent une augmentation ou une perte de l'activité des récepteurs. Exemples d'acides aminés hautement conservés : Asn1.50, Asp2.50, Trp4.50 and Pro7.50 Source : Venkatakrishnan et al. (2013) En termes de positionnement spatial au sein du récepteur, les contacts tertiaires consensus inter-TM sont en grande partie localisés au centre et du côté cytoplasmique de l'ensemble TM et regroupés en particulier aux interfaces TM1-TM2, TM3-TM4, TM3-TM5 et TM3-TM6-TM7. |
c. Particularités de la région intracellulaire Les résidus de la région intracellulaire et les extrémités cytoplasmiques des TM lient des effecteurs de signalisation tels que les protéines G, les kinases de GPCR et les arrestines. Dans les structures disponibles :
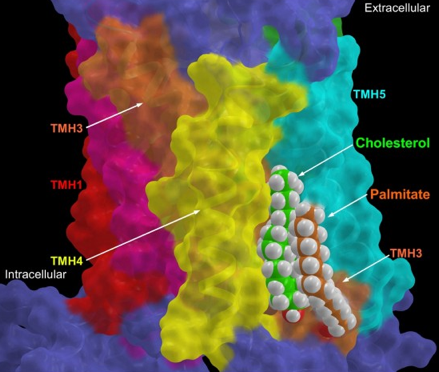
En plus des ICL, une hélice amphipathique courte (H8), contenant généralement trois tours d'hélice α et des sites de palmitoylation à son extrémité C-terminale, est présente dans plusieurs structures de GPCR de la classe A. Source : Zheng et al., 2012 C3.55 est le site de palmitoylation de OPRM1 (récepteur des μ-opioides). La palmitoylation du récepteur facilite l'association du cholestérol pour former un complexe de signalisation [cholestérol-palmitoyl-OPRM1]. L'interaction cholestérol-palmitoyl contribue au mécanisme de signalisation de OPRM1 en facilitant son homodimérisation et son couplage avec les protéines Gαi2. ICL3 et l'extrémité C-terminale sont des régions longues et variables. Elles sont probablement intrinsèquement non structurées chez beaucoup de GPCR. Les régions intrinsèquement non structurées exposent des motifs peptidiques linéaires qui reconnaissent des partenaires spécifiques, ce qui régule leur fixation et leur fonction. Diverses kinases et le domaine tyrosine kinase du récepteur de l'insuline peuvent phosphoryler les régions cytoplasmiques des β-adréno-récepteurs. Les différentes formes phosphorylées de ces récepteurs établissent des interactions différentes avec la β-arrestine, ce qui influence l'activité du récepteur et l'internalisation à partir de la membrane. |
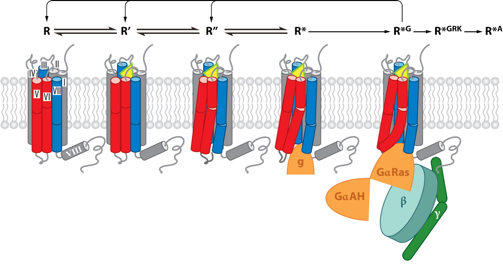
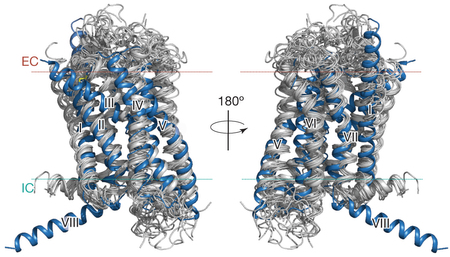
d. Intermédiaires structuraux de l'activation d'un RCPG Des intermédiaires clé du mécanisme d'activation des RCPG ont été caractérisés par cristallographie (figure ci-dessous).
Source : Katritch et al., 2013
|
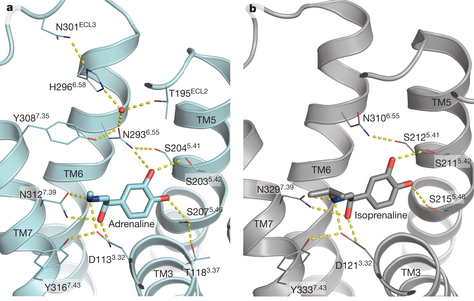
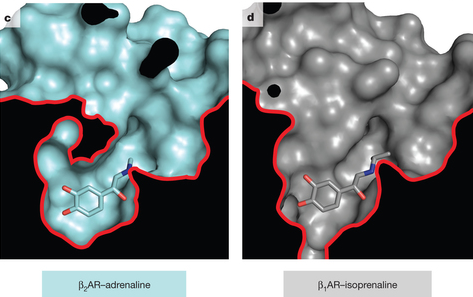
5. Comparaison des conformations active et inactive d'un RCPG modèle L'équipe de Brian Kobilka* a conçu un fragment d'anticorps de haute affinité qui stabilise la conformation active de β2AR (β2-adrénorécepteur). Les structures de β2AR lié à 3 agonistes chimiquement distincts ont été déterminées :
Cette technique a permis de comparer pour la première fois, les structures d'un récepteur adrénergique liés à des catécholamines dans ses conformations actives et inactives. [*Robert Lefkowitz et Brian Kobilka ont reçu le prix Nobel de Chimie en 2012 pour leurs travaux sur les RCPG]. Les résultats indiquent que :
Source : Ring et al., 2013
Source : Ring et al., 2013 L'utilisation de 2 nano-anticorps (Nb80 qui se fixe à β2AR et Nb37 qui se fixe à la forme Gs dépourvue du nucléotide) ont apporté des preuves expérimentales qui renforcent l'hypothèse que les RCPG localisés dans les endosomes sont encore actifs. |
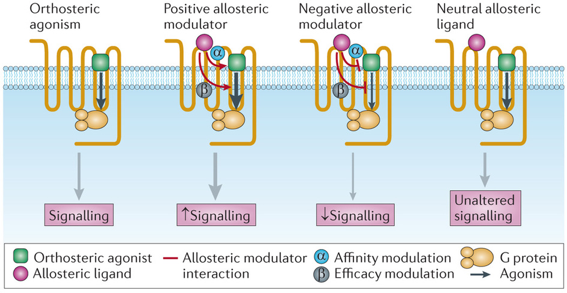
RCPG et site de fixation orthostérique Les agonistes orthostériques se fixent à un RCPG et induisent un changement conformationnel du RCPG ce qui déclenche la cascade de signalisation en aval.
Les ligands allostériques qui n'ont aucun effet sur l'affinité et/ou l'efficacité médiée par l'agoniste orthostérique sont appelés ligands allostériques neutres. Source : Wootten et al., 2013 Les flèches rouges indiquent les interactions allostériques entre le modulateur et le ligand orthostérique. Les flèches noires indiquent les interactions allostériques entre les sites de fixation du ligand et le site de fixation de l'effecteur au sein du RCPG, qui entraînent l'activation des voies de signalisation, appelé aussi "agonisme orthostérique". |
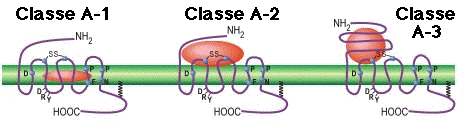
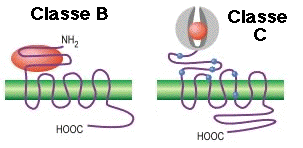
6. Comparaison des RCPG de la classe A et de la classe B Le récepteur du glucagon (GCGR) est l'un des 15 membres des RCPG de la classe B ("secretin-like"). La fixation du glucagon au récepteur au GCGR libère le glucose du foie pendant le jeûne. Le GCGR joue donc un rôle important dans l'homéostasie du glucose. Exemples de récepteurs de la classe B :
Les principales différences entre le GCGR et les RCPG de la classe A sont :
Source : Siu et al. (2013) - GCGR en bleu et RCPG de la classe A en gris. Caractéristiques structurales des RCPG de la classe B
Remarque : la nomenclature en vigueur pour les RCPG de la classe B est celle de Wootten et al. (2013). |
| 8. Liens Internet et références bibliographiques |
|
Base de données : "GPCRDB: Information system for G protein-coupled receptors (GPCRs)" Base de données : "Orientation of Proteins in Membranes" Nobel Prize in Chemistry 2012 : Robert J. Lefkowitz and Brian K. Kobilka |
|
|
Ballesteros & Weinstein (1995) "Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors" Methods neurosciences 25, 366 - 428 Hermans and Challiss (2001) "Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: prototypic family C G-protein-coupled receptors" Biochemical J. 359, 465 - 484 Karlin A. (2002) "Emerging structure of the Nicotinic Acetylcholine receptors" Nature Rev. Neuroscience 3, 102 - 114 Rasmussen et al. (2011) "Crystal structure of the β2 adrenergic receptor–Gs protein complex" Nature 477, 549 - 555 Chandra et al. (2013) "Multidomain integration in the structure of the HNF-4α nuclear receptor complex" Nature |
|
Gurevitch & Gurevitch (2006) "The structural basis of arrestin-mediated regulation of G-protein-coupled receptors" Pharmacol. Ther. 110, 465 - 502 Smrcka, AV. (2009) "G protein βγ subunits: Central mediators of G protein-coupled receptor signaling" Cell Mol. Life Sci. 65, 2191 - 2214 Pelé et al. (2011) "Multidimensional Scaling Reveals the Main Evolutionary Pathways of Class A G-Protein-Coupled Receptors" PLoS ONE 6(4): e19094 Zheng et al. (2012) "Palmitoylation and membrane cholesterol stabilize μ-opioid receptor homodimerization and G protein coupling" BMC Cell Biol. 13, 6 Lin et al. (2013) "A pharmacological organization of G protein–coupled receptors" Nature Methods 10, 140 - 146 |
|
Wheatley et al. (2012) "Lifting the lid on GPCRs: the role of extracellular loops" Br. J. Pharmacol. 165, 1688 - 1703 Venkatakrishnan et al. (2013) "Molecular signatures of G-protein-coupled receptors" Nature 494, 185 - 194 Ring et al. (2013) "Adrenaline-activated structure of β2-adrenoceptor stabilized by an engineered nanobody" Nature, doi:10.1038/nature12572 Siu et al. (2013) "Structure of the human glucagon class B G-protein-coupled receptor" Nature 499, 444 - 449 Katritch et al. (2013) "Structure-Function of the G Protein–Coupled Receptor Superfamily" Ann. Rev. Pharmacol. Toxicol. 53, 531 - 556 Hauser et al. (2021) "GPCR activation mechanisms across classes and macro/microscales" Nat. Struct. Mol. Biol. 28, 879 - 888 |
![]()