| L'isoprénylation, la myristoylation, la palmitoylation, la glypiation |
| Tweet |
|
|
1. Introduction 2. L'isoprénylation : substrats (farnesyl ou géranylgéranyl) et réaction 3. La myristoylation : substrats (acide myristique) et réaction 4. La palmitoylation |
5. La glypiation
6. Liens Internet et références bibliographiques |
|
1. Introduction Les modifications les plus courantes des protéines par des lipides sont l'isoprénylation, la N-myristoylation, la palmitoylation (ou S-acylation) et la glypiation.
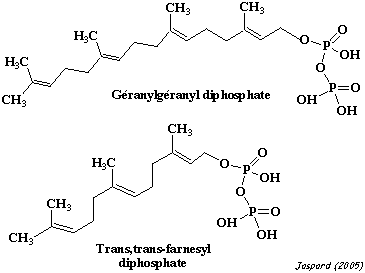
Les groupements farnesyl ou géranylgéranyl, sont des produits de la voie métabolique de l'HMG-CoA réductase. |
| Exemples d'acides gras | |||||
| Carbone | Doubles liaisons | Position des doubles liaisons | Nom commun | Nom IUPAC | Point de fusion |
| 12 | 0 | C12:0 | laurate (acide laurique) |
dodécanoate (acide dodécanoique) |
43°C |
| 14 | 0 | C14:0 | myristate | tétradécanoate | 52°C |
| 16 | 0 | C16:0 | palmitate | hexadécanoate | 63°C |
| 18 | 0 | C18:0 | stéarate | octadécanoate | 70°C |
| 20 | 0 | C20:0 | arachidate | icosanoate | 75°C |
| 22 | 0 | C22:0 | béhénate | docosanoate | 81°C |
| 24 | 0 | C24:0 | lignocérate | tétracosanoate | 84°C |
| 16 | 1 | C16:1 Δ9 | palmitoléate | cis-Δ9-hexadécénoate | - 0,5°C |
| 18 | 1 (végétaux) | C18:1 Δ9 (acide gras ω-9) | oléate | cis-Δ9-octadécénoate | 13°C |
| 18 | 2 (végétaux) | C18:2 Δ9,12 (acide gras ω-6) | linoléate | cis,cis-Δ9,12-octadécadiénoate | - 9°C |
| 18 | 3 (végétaux) | C18:3 Δ9,12,15 (acide gras ω-3) | linolénate | all-cis-Δ9,12,15-octadécatriénoate | - 17°C |
| 18 | 4 (végétaux) | C18:4 Δ6,9,12,15 (acide gras ω-3) | stéaridonate | all-cis-Δ6,9,12,15-octadécatétraénoate | --- |
| 20 | 4 (animaux) | C20:4 Δ5,8,11,14 (acide gras ω-6) | arachidonate | all-cis-Δ5,8,11,14-icosatétraénoate | - 49°C |
| 20 | 5 (poisson) | C20:5 Δ5,8,11,14,17(acide gras ω-3) | eicosapentaénoate | all-cis-Δ5,8,11,14,17-eicosapentaénoate | --- |
| 22 | 5 (animaux) |
C22:5 Δ4,7,10,13,16 (acide gras ω-6) C22:5 Δ7,10,13,16,19 (acide gras ω-3) |
acide osbond clupanodonate |
all-cis-Δ4,7,10,13,16-docosapentaénoate all-cis-Δ7,10,13,16,19-docosapentaénoate |
--- |
| 22 | 6 (animaux) | C22:6 Δ4,7,10,13,16,19 (acide gras ω-3) | cervonate | all-cis-Δ4,7,10,13,16,19-docosahexaenoate | - 44°C |
| 24 | 6 | C24:6 Δ6,9,12,15,18,21 (acide gras ω-3) | nisinate | all-cis-Δ6,9,12,15,18,21-tétracosahexaénoate | --- |
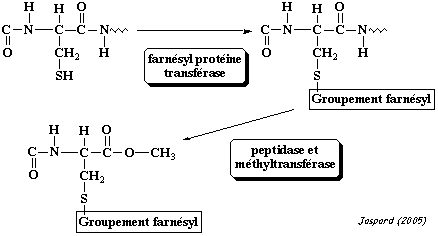
2. L'isoprénylation : substrats et réaction L'isoprénylation (ou prénylation) est la modification qui ajoute un lipide isoprénoide farnesyl ou géranylgéranyl, via une liaison thioether, sur une cystéine en position C-terminale ou proche de cette extrémité.
La protéine est ensuite clivée juste après la cystéine modifée et un groupement méthyle y est ajouté en position C-terminale. Les groupements farnesyl (15 carbones) et géranylgéranyl (20 carbones) sont des poly-isoprenes insaturés. Ils dérivent du mévalonate. Chez l'homme il existe 3 protéine prényltransferases : la farnesyl-transférase (FT) et les géranylgéranyl-transférase 1 et 2 (GGT1 et GGT2). La réaction catalysée par la GGT2 met en jeu 2 molécules de géranylgéranyl diphosphate, au lieu de 1 pour FT et GGT1. Chacune de ces protéines contient une sous-unité α et une sous-unité β. Les sous-unités α de FT et de GGT1 sont codées par le même gène (FNTA).
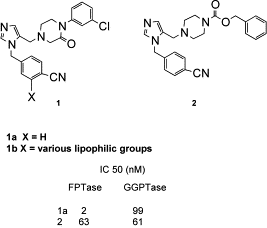
La séquence consensus d'isoprénylation est : "Ca1a2X" où C est la cystéine modifiée. Figure ci-contre : structure d'un inhibiteur de la FT et de la GGT1.
Source : Tucker et al. (2002)
|
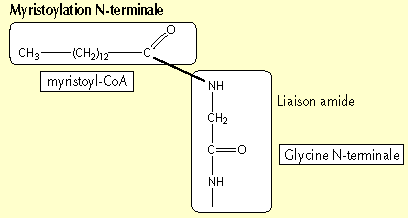
3. La myristoylation : substrats et réaction La N-myristoylation est la modification qui ajoute un acide myristique (activé sous forme de myristoyl-CoA) sur une glycine en position N-terminale. La méthionine N-terminale issu du codon d'initiation de la traduction est au préalable clivée.
Source : Rioux & Legrand (2001) L'acide myristique (C14:0) est présent en faible quantité dans les tissus animaux. Il ne représente que 0,6 % des acides gras totaux du foie de souris. La O-acylation : liaison oxyester entre un acide gras et une sérine interne. C'est une modification des protéines membranaires virales : par exemple, la protéine gag du virus HIV est myristoylée et sans cette modification, le virus ne peut pas produire de particules infectieuses. La rhodopsine bien que possèdant 7 domaines transmembranaires, utilise une ancre S-palmitoylée pour attacher l'extrémité C-terminale cytoplasmique à la membrane plasmique.
|
|
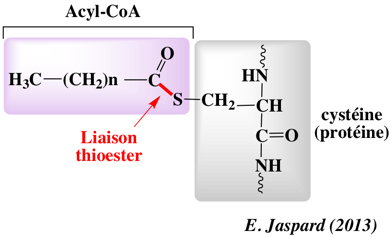
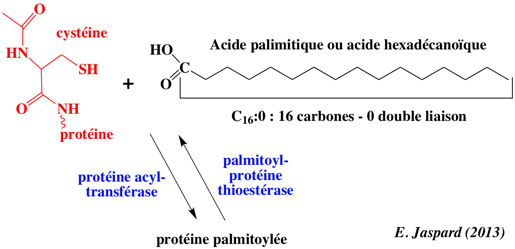
La S-palmitoylation (ou S-acylation) est une modification post-traductionnelle qui ajoute, dans la grande majorité des cas, un acide palmitique (acide gras saturé en C16) sur 1 à 4 cystéine(s) d'une protéine membranaire, via une liaison thioesther. palmitoyl-CoA + [protéine]-L-cysteine <=> [protéine]-S-palmitoyl-L-cysteine + CoA
Les protéines sont souvent modifiées de manière séquentielle par des lipides différents :
La palmitoylation a lieu dans le cytoplasme et n'utilise pas les voies de sécrétion du réticulum endoplasmique. La palmitoylation augmente l'hydrophobicité de la protéine et donc son aptitude à s'associer aux membranes. La palmitoylation est essentielle pour le repliement des protéines membranaires impliquées dans la signalisation et le trafic intracellulaires comme les kinases de la famille Src, les GTPases de la famille Ras, les protéines G et les récepteurs couplés aux protéines G . La palmitoylation est aussi impliquée dans le métabolisme des lipoprotéines, l'apoptose et d'autres processus cellulaires. |
La S-palmitoylation est catalysée par les palmitoyl S-acyltransférases qui ont fixées à la membrane. A l'inverse de la prénylation et de la myristoylation, la palmitoylation est en général réversible du fait de la nature chimique de la liaison thioester établie entre l'acide gras et l'acide aminé. La dé-palmitoylation est catalysée par les palmitoyl-protéines thioesterases (EC 3.1.2.22).
Ce processus de[palmitoylation - dépalmitoylation] est la seule modification par des qui soit réversible. Cette dynamique d'états modifiés ou pas est un élément clé de la régulation des protéines membranaires palmitoylées, au même titre que la [phosphorylation (kinases) - déphosphorylation (phosphatases)] des protéines. Palmitoyl-protéines thioesterases : PDB 3GRO. Les palmitoyl S-acyltransférases (EC 2.3.1.225) contiennent un domaine conservé riche en cystéine avec un motif [DHHC] nécessaire à l'activité enzymatique. C'est la raison pour laquelle on les appelle aussi les protéines DHHC. Ce motif est lui-même inclus dans une région plus grande d'environ cinquante acides aminés très conservés, dont le le motif général est : [C-x(2)-C-x(9)-HC-x(2)-C-x(4)-DHHC-x(5)-C-x(4)-N-x(3)-F] Elles protéines S-acyltransférases s'auto-acylent. Les palmitoyl acyltransférases DHHC5, DHHC6 et DHHC8 sont S-acylées sur 3 résidus Cys au niveau d'un motif [CCX(7-13)C (S/T)] situé en aval du domaine DHHC conservé. Cette auto-acylation est un mécanisme qui régule probablement la spécificité et/ou l'activité des acyltransférases. |
|
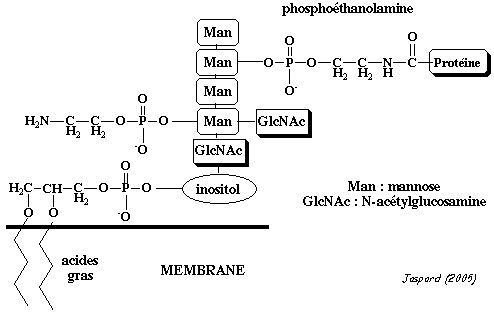
5. La glypiation 5a. Structure du substrat GPI et structure du site de fixation du GPI La glypiation est la modification qui ajoute un groupement glycosyl-phosphatidylinositol ou GPI ou ancre GPI sur un acide aminé en position C-terminale de protéines fixées à la membrane du réticulum endoplasmique. La protéine est fixée à la membrane avant glypiation et le reste après, mais par 2 entités structurales différentes :
Chez Saccharomyces cerevisiae, l'ancrage GPI est essentiel pour la croissance : sur environ 6200 phases de lecture ouvertes ressencées sur son génome, 60 codent des protéines à ancre GPI. Du fait qu'elles contiennent des phospholipides, les structures des ancres GPI confèrent une grande mobilité aux protéines membranaires ainsi ancrées. Une autre caractéristique des ancrages GPI est l'activation potentielle des protéines de la membrane ainsi ancrées, qui peuvent être relarguées par un mécanisme de signalisation impliquant une phospholipase C spécifique (EC 3.1.4.3). L'ancre GPI est pré-assemblée dans la membrane.
Elle est composée :
L'acide myristique intervient parfois dans la glypiation. Les ancrages GPI par des formes dipalmitoylées sont les plus courants mais des ancrages par des formes dimyristoylées sont également observés. |
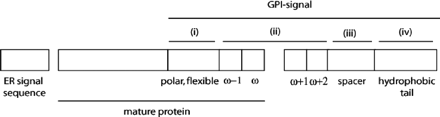
Le schéma ci-dessous indique les 2 séquences signal nécessaires pour un ancrage GPI :
Source : Eisenhaber et al. (2003) Cette dernière contient 4 régions (notées i à iv - figure ci-dessus) :
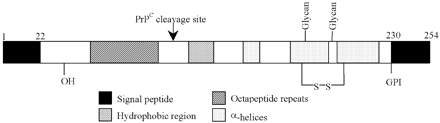
La forme mature de la protéine après les modifications liées à la glypiation est précisée sur la figure. Figure ci-dessous : structure shématique de la protéine prion PrP. Sont indiqués :
Source : Gill et al. (2000) |
| composant | acides aminés | rôle |
| GPI8 | 411 | précurseur de la transamidase GPI proprement dite (EC 3.XXX - peptidase C13) |
| GAB1 | 394 | impliqué dans la reconnaissance du peptide signal d'ancrage du GPI ou dans la reconnaissance de la partie lipidique du GPI |
| GPI16 | 602 | impliqué dans le transfert du GPI sur la protéine substrat |
| GPI17 | 534 | |
| GAA1 | 614 | recquise pour l'étape terminale de l'ancrage du GPI sur la protéine substrat affecte l'endocytose |
On peut noter qu'il existe un autre mécanisme enzymatique : les acides aminés du peptide signal C-terminal sont relargués par une carboxypeptidase, suivi de la fixation du GPI par une transferase. 5d. Réaction de pré-assemblage du GPI (2è étape) Voir la voie de biosynthèse : KEGG La seconde étape de la synthèse de l'ancre GPI est catalysée chez tous les eucaryotes par la N-acétylglucosaminylphosphatidylinositol désacétylase (GPI12 - EC 3.5.1.89). Elle est située dans la membrane du réticulum endoplasmique. Réaction : 6-(N-acetyl-D-glucosaminyl)-1-phosphatidyl-1D-myo-inositol + H2O <==> 6-(alpha-D-glucosaminyl)-1-phosphatidyl-1D-myo-inositol + acétate |
| Récapitulatif des principales caractéristiques des 4 classes majeures d'acylation des protéines
Source : S. Patterson (2002) |
||||
| Class of protein acylation | N-Myristoylation | S-Palmitoylation | Prenylation | Glypiation |
| Type of event | Cotranslational | Posttranslational | Cotranslational | Cotranslational and posttranslational |
| Group and bond | Amide-linked fatty acid | Thioester-linked fatty acid | Thioether-linked isoprenoid | Polysaccharide-linked phosphatidylinositol |
| Location of modification | N-terminal glycine | Predominantly cysteine anywhere in the protein | C-terminal cysteine | C-terminal amino acid after signal cleavage |
| Reversibility | Stably bonded | Fatty acid turns over | Stably bonded | Stably bonded with fatty acid remodelling |
| Association with protein synthesis | One step, NMT | Not known | Two-step, Ftase, GGtase | Complex synthesis |
| Type of association | With proteins | Membrane insertion | With proteins, maybe membrane insertion | Membrane insertion |
| Type of proteins modified | Intracellular proteins | Intracellular or transmembrane proteins | Intracellular proteins | Cell surface proteins |
| Effect of modification | Reversible inter- or intra- protein associations | Proteins localize to rafts and polarized cell compartments | Proteins associate reversibly with various intracellular membranes/proteins | Proteins localize to rafts in apical or axonal membrane domains |
| Abbréviations : NMT, N-myristoyl-transférase - Ftase, farnesyl-transférase - GGtase, géranylgéranyl-transférase | ||||
| 6. Liens Internet et références bibliographiques |
|
"PrePS - Prenylation Prediction Suite" : Site web pour la prédicion de sites de farnesylation (CaaX), de géeranylgéranylation (CaaX) et de géranylgéranylation (Rab) "Proteolipids Structure, Occurrence And Biology" Myristoylation : "Myristoylator" - Site web pour la prédicion de site de N-myristoylation (réseau de neurones) Glypiation : "Big-PI Plant Predictor" - Site web pour la prédiction de site de modification par ancrage GPI chez les plante PRENbase : Base de données spécifiques des protéines prénylées [farnesylation (CaaX), de géeranylgéranylation (CaaX) et de géranylgéranylation (Rab)] |
|
|
Gill et al. (2000) "Post-translational hydroxylation at the N-terminus of the prion protein reveals presence of PPII structure in vivo" EMBO J. 19, 5324 - 5331 S.Patterson (2002) "Posttranslational protein S-palmitoylation and the compartmentalization of signaling molecules in neurons" Biol. Res. 35, 139 - 150 Tucker et al. (2002) "The synthesis and biological evaluation of a series of potent dual inhibitors of farnesyl and geranyl-Geranyl protein transferases", Bioorganic & Medicinal Chemistry Letters 12, 2027 - 2030 Eisenhaber et al. (2003) "Glycosylphosphatidylinositol Lipid Anchoring of Plant Proteins. Sensitive Prediction from Sequence- and Genome-Wide Studies for Arabidopsis and Rice" Plant Physiol. 133, 1691 - 1701 |
![]()