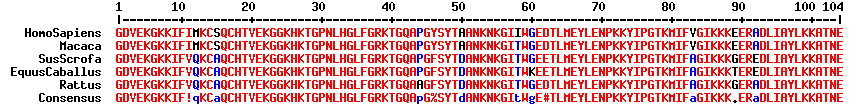| Le cytochrome c, la chaîne respiratoire et l'apoptose |
| Tweet |
|
|
1. Les cytochromes 2. Le cytochrome c 3. Phylogénie et historique de la photosynthèse et de la respiration 4. Implication du cytochrome c dans l'apoptose |
5. Les 2 voies de l'apoptose 6. Signalisation de l'apoptose 7. Les caspases 8. Liens Internet et références bibliographiques |
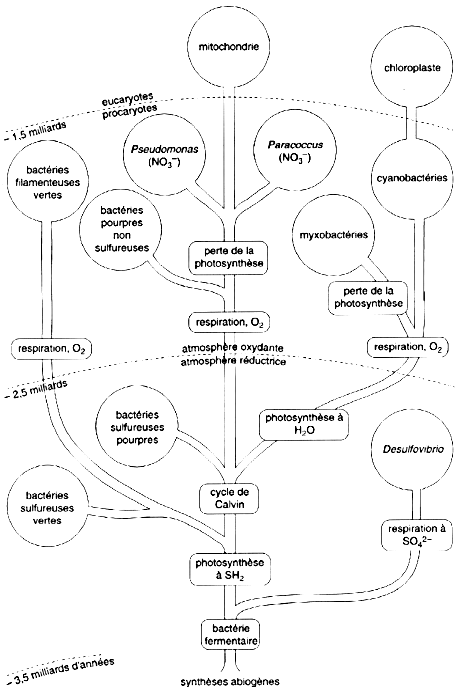
1. Les cytochromes Le nom de cytochrome a été proposé par D. Keilin en 1925. Ce sont des protéines qui contiennent un ou plusieurs groupements hème : un atome de fer à l'état ferreux (FeII) ou ferrique (FeIII) chélaté par un groupe tétrapyrrole. L'état d'oxydation Fe(IV) est trouvé dans certains systèmes enzymatiques comme intermédiaire catalytique. Un hémochrome est défini comme un composé hème de spin faible au sein duquel la 5ème et la 6ème position de coordination sont occupées par des ligands à fort champs par rapport à l'état d'oxydation du fer. Il existe 4 groupes de cytochromes :
La classification est aussi liée à la plus faible longueur d'onde d'absorbance du groupement hème (bande d'absorbance de la pyridine ferro-hémochrome en solution alcaline) lors de la réduction :
Il existe d'autres transporteurs d'électrons dans les cellules qui contiennent d'autres types de groupements prosthétiques. Exemples : la ferrédoxine (fer et soufre), l'azurine (cuivre), la plastocyanine. |
|
Ces sont les premières séquences de protéines qui ont été intégrées dans la base de données Swiss-Prot/Uniprot.
Le cytochrome c est une petite protéine trés soluble (70 à 120 acides aminés selon la classe - voir ci-dessous / 105 acides aminés chez l'homme - masse molaire = 11,8 kDa), associée à la membrane interne des mitochondries. Le génome de Geobacter sulfurreducens code pour plus de 100 cytochromes c (!) qui acceptent des donneurs et des accepteurs d'électrons trés divers ce qui laisse supposer de grandes aptitudes d'adaptation via de multiples voies métaboliques. Il existe 4 classes de cytochromes c (Ambler, 1991) :
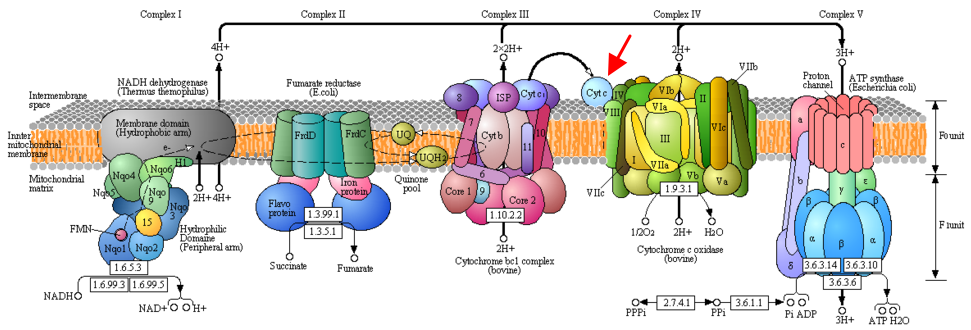
Le cytochrome c est un composant essentiel de la chaîne de transport des électrons (figure ci-dessous).
Source : KEGG - "Oxidative phosphorylation" La figure originale est interactive et permet d'obtenir un trés grand nombre d'informations concernant les gènes, les protéines et autres pour les divers complexes. Le complexe III de la chaîne respiratoire Le cytochrome c est un transporteur mobile de la chaîne respiratoire. Il est constitué entre autre d'une protéine Fe - S, d'un cytochrome b qui porte les groupes hème b560 et b566 et du cytochrome c1. Les électrons sont cédés par le complexe III au cytochrome c (attention : différent du cytochrome c1). Puis les 4 électrons sont transferés un à un du cytochrome c au complexe IV. L'ordre de transfert des électrons est : cytochrome c ---> atome de cuivre A ---> hèmes cytochrome a ---> [hème cytochrome a3 / atome de cuivre B]. Le tracé des électrons n'a été résolu que quand un trajet circulaire, appelé cycle Q, fût suggéré par Peter Mitchell puis détaillé entre autres par Bernard Trumpower. Le coenzyme Q et le coenzyme QH2 diffusent d'une face à l'autre de la membrane mitochondriale interne et l'ensemble des deux hèmes du cytochrome b occupe toute l'épaisseur de cette membrane. Bilan du cycle Q : coenzyme QH2+ 2 cyt coxydé + 2 H+ matrice ---> coenzyme Q + 2 cyt creduit + 2 H+espace intermembranaire La variation d'énergie libre de Gibbs liée au parcours des électrons au sein du complexe III permet l'expulsion de 2 x 2 protons vers l'espace intermembranaire. Le complexe III contribue à la force protons motrice. |
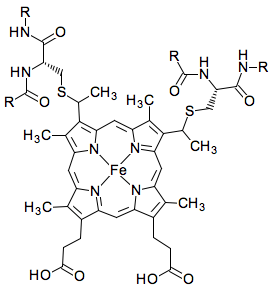
L'hème contient un atome de fer au sein d'un noyau porphyrine.
Source : KEGG - "Oxidative phosphorylation" L'atome de fer est lié à 4 atomes d'azote du noyau porphyrine. Seul l'hème du cytochrome c est covalemment lié à des résidus cystéine via une liaison thioether. Les groupements hèmes des groupes a, b et c de cytochrome diffèrent légèrement au niveau des substituants du noyau porphyrine. Par exemple, l'hème du cytochrome a possède une longue chaîne farnesyle qui inclue 3 unités isoprènoides. Schéma de la fixation de l'hème au cytochrome c mitochondrial Les groupement vinyle de l'hème sont saturés par l'addition de groupements thiols de Cys inclues dans un motif C14XXC17H18.
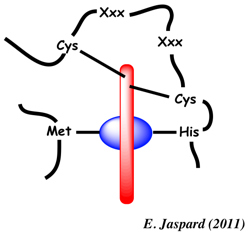
Adapté de Allen et al. (2008) Les Cys établissent donc des liaisons covalentes entre l'hème (noyau porphyrine en bleu) et le cytochrome c et l'atome d'azote de His ligande l'atome de fer (en bleu) de l'hème. La 6ème liaison de coordination à l'atome de fer est fournie par Met80 (chez les Eucaryotes) située vers l'extrémité C-terminal. Selon les règnes, on trouve d'autres acides aminés à la place de la Met : His chez les bactéries, Cys, un acide aminé N-terminal, Asn, Lys, voire un site de coordination libre. Voir une étude des acides aminés invariants des cytochromes. Le cytochrome c et d'autres cytochromes sont parmi les protéines qui ont été le plus étudiées sur le plan structural (grande facilité d'obtention, stabilité de la structure se prêtant à la cristallisation, petit nombre d'acides aminés, ...). Il existe plus d'une centaine de structures dans la base de données PDB. Ci-dessous, cytochrome c du thon.
|
|
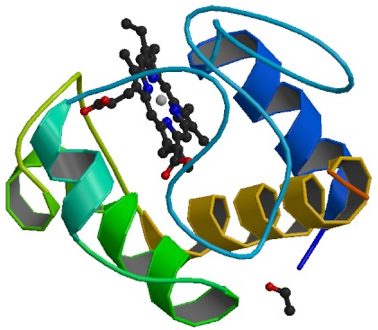
Visualisation du cytochrome c du cheval à une résolution de 1,90 Å. Code PDB : 1HRC
|
|
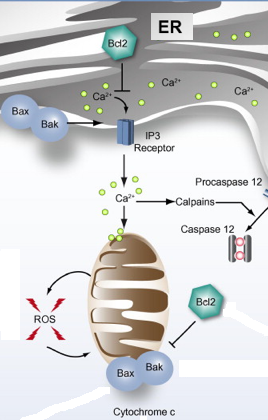
4. Implication du cytochrome c dans l'apoptose Certaines protéines pro-apoptotiques de la famille Bcl-2 ("B-cell lymphoma 2") telles que Bax et Bak sont localisées sur la membrane du réticulum endoplasmique (RE) et interagissent avec le domaine cytosolique d'IRE-1 ("Inositol-Requiring Enzyme-1"). Elles régulent l'homéostasie calcique.
Source : Malhi & Kaufman (2011) Remarque : Bcl-2 est à la fois le nom de la famille et le nom de protéines anti-apoptotiques. Le RE peut relarguer du calcium intracellulaire via les canaux sous contrôle du récepteur de l'inositol 1,4,5-triphosphate (IP3R). Le relarguage du calcium du RE peut activer des calpaïnes, qui peuvent à leur tour activer par protéolyse la caspase 12, un médiateur de l'apoptose. L'afflux de calcium dans la mitochondrie altère la perméabilité de la membrane mitochondriale ce qui induit un relarguage du cytochrome c dans le cytosol via des pores de transition de perméabilité (PT).
Source : RCDRG Le pore PT est un canal constitué de :
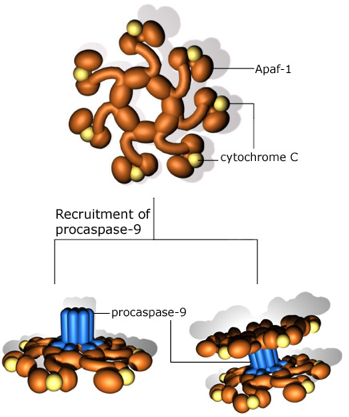
La fixation du cytochrome c à la protéine Apaf-1 ("Apoptotic peptidase activating factor") entraîne l'activation de la caspase 9 qui accélère l'apoptose en activant d'autres caspases ("aspartic-acid-specific cystein proteases"). |
|
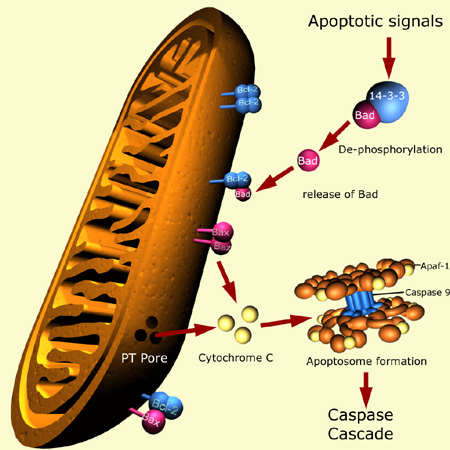
Entre 50 milliards et 70 milliards de cellules meurent chaque jour par apoptose chez l'humain adulte. a. La voie intrinsèque Cette voie est régulée par les membres de la famille Bcl-2 et la fonction des effecteurs est médiée par le relarguage de protéines comme le cytochrome c après perméabilisation de la membrane mitochondriale externe, comme décrit ci-dessus. Le cytochrome c, la protéine Apaf-1, la pro-caspase 9 et l'ATP forment un complexe multi-protéique appelé l'apoptosome (figure ci-dessous).
Source : RCDRG Apaf-1 contient :
La caspase 9 est activée et elle active à son tour la caspase 3. Ces caspases hydrolysent des protéines clé comme celles du cytosquelette, ce qui entraîne un changement typique de la morphologie des cellules observée lors de l'apoptose.
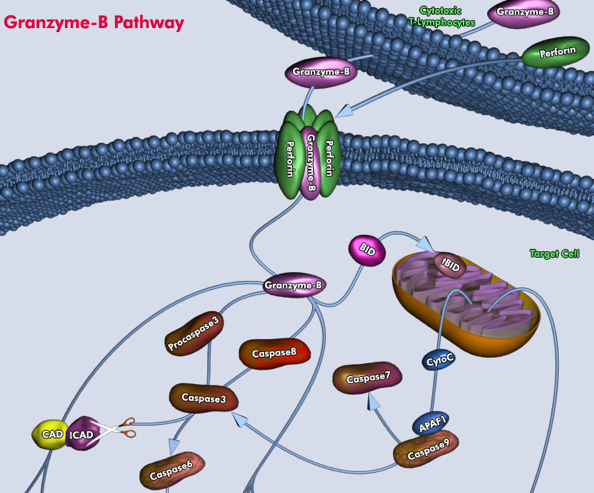
Source : RCDRG b. La voie extrinsèque Elle est appelée aussi la voie du récepteur de mort cellulaire ("death receptor pathway"), est déclenchée par la fixation de molécules à des récepteurs de l'apoptose qui appartiennent à la famille "tumor necrosis factor" ou TNF. Ces récepteurs contiennent un domaine intracellulaire de signalisation de mort cellulaire qui recrute et active des caspases initiatrices telles que la caspase 8 ou la caspase 10. Il s'en suit l'activation de caspases effectrices telles que la caspase 3, la caspase 6 ou la caspase 7. La voie extrinsèque a lieu indépendamment de la famille Bcl-2. Les 2 voies ont donc rarement lieu ensemble, sauf dans les hépatocytes, où l'hydrolyse par la caspase 8 médie l'activation de la protéine "BH3-only" Bid , un membre pro-apoptotique de la famille Bcl-2. Bid est activée par protéolyse par la caspase-8 au niveau de sa région intrinsèquement désordonnées ("Intrinsically disordered regions").
Source : Qiagen En dehors de ceux médiés par l'activation des récepteurs de mort cellulaire, il existe des mécanismes qui déclenchent aussi une cascade d'activation de caspases. Par exemple, la granzyme B relarguée dans les cellules par les lymphocytes T cytotoxiques, peut activer directement les caspases 3, 7, 8 et 10. La granzyme B est une protéase à sérine. |
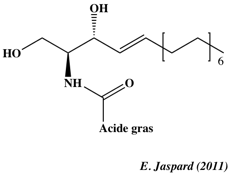
6. Signalisation de l'apoptose a. Via les récepteurs de mort cellulaire Les récepteurs de mort cellulaire sont situées à la surface des cellules. Ils transmettent les signaux apoptotiques qui sont initiés par des ligands spécifiques tels que FasL ("Fas Ligand"), TNF α et TRAIL. La fixation du ligand aux récepteurs de mort cellulaire peut générer des céramides produits par la sphingomyelinase acide (ou sphingomyeline phosphodiestérase - EC 3.1.4.12) à partir de la sphingomyeline. Les céramides (figure ci-dessous) sont une famille de sphingolipides , composés d'une sphingosine et d'un acide gras.
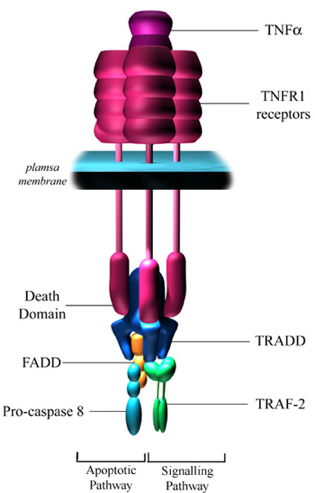
Les sphingolipides sont en grande quantité dans les membranes des cellules. Cette formation importante de céramides entraîne la fusion de lipides qui induit un regroupement d'un très grand nombre de récepteurs de mort cellulaire, ce qui a pour conséquence d'amplifier le signal de la mort cellulaire. En l'absence de regroupement de récepteurs de mort cellulaire, certaines cellules comme les lymphocytes sont malgré tout capables de déclencher l'apoptose. Mais dans la plupart des cas, ce mode d'amplification est nécessaire à la pleine activation de la réponse apoptotique. La fixation du ligand au récepteur de mort cellulaire induit un changement de conformation du domaine intracellulaire du récepteur. Ce changement révèle un domaine de mort cellulaire qui lui permet de recruter de nombreuses protéines apoptotiques. Ce complexe protéique est appelé DISC ("Death Inducing Signalling Complex"). L'étape suivante est le recrutement d'une caspase (typiquement la caspase 8) par DISC, caspase qui est activée et qui initie l'apoptose. b. Via les récepteurs TNF α ("tumor necrosis factor α" ou cachexine) TNF α est une cytokine (glycoprotéine - 185 acides aminés) produite par les lymphocytes T et les macrophages activés en reponse à l'infection et aux processus inflammatoires L'activation du récepteur de TNF - le récepteur TNFR1 a de nombreux effets. La fixation de TNF α à TNFR1 induit la trimèrisation du récepteur et le regroupement de domaines intracellulaires de mort cellulaire.
Source : RCDRG Une molécule adaptatrice appelée TRADD ("TNF-RI-associated with death domain") peut alors s'y fixer via les interactions entre les domaines de mort cellulaire. TRADD peut recruter un grand nombre de protéines différentes au récepteur activé. Le recrutement de TRAF2 ("TNF-associated factor 2") peut conduire à l'activation des voies de signalisation NF-kB et JNK ("c-Jun N-terminal kinase"). JNK a des effets pro-apoptotiques : activation induite par phosphorylation de Bim et inactivation des protéines Bcl2. TRADD peut aussi s'associer à FADD ("Fas associated protein with death domain"), ce qui conduit à l'induction de l'apoptose via le recrutement et l'hydrolyse de la pro-caspase 8. |
7. Les caspases ("Cysteine-dependent ASPartyl-specific proteASES") En 2011, on dénombre 15 caspases. Elles ne sont pas toutes impliquées dans l'apoptose. Par exemple, la caspase 1 ("interleukin-1-beta converting enzyme" - ICE) est impliquée dans les processus inflammatoires. Il existe 2 types de caspases apoptotiques :
Les caspases qui contiennent un domaine CARD ("CAspase Recruitment Domain") sont soit :
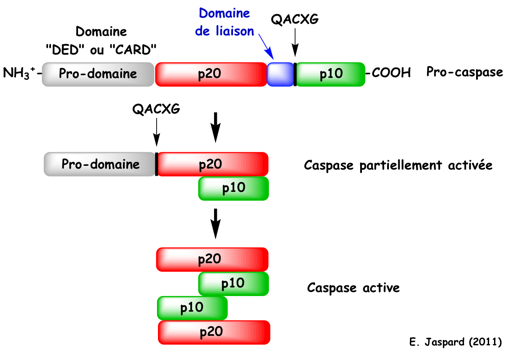
Les caspases appartiennent à la famille des protéases à cystéine et sont biosynthétisées sous la forme de précurseurs inactifs (ou zymogènes) : les pro-caspases. Ces zymogènes sont activés par coupure protéolytique par des caspases ou par auto-activation. Les caspases ont une structure très conservée :
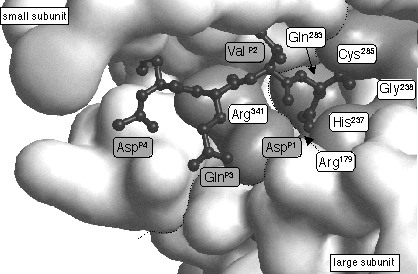
L'activation du zymogène en caspase active s'effectue par 2 coupures protéolytiques successives au niveau d'une liaison Asp-X (Asp en position P1 - voir la figure ci-dessous). La première coupure protéolytique libére la petite sous-unité et la seconde libére le pro-domaine. La caspase active est un tétramère composé de deux p20 et deux p10. Le site catalytique du domaine p20 des caspases est très conservé : motif QACXG où C est la Cys active et X = Gly, Gln ou Arg.
Source : Denault & Salvesen (2002) Les caspases sont trés sélectives (voir des exemples de cibles des caspases). On note qu'un grand nombre de protéines kinases sont protéolysées pendant l'apoptose. Structures 3D : caspase 3 / PDB 1PAU - caspase 6 / PDB 3OD5 - pro-caspase 7 / PDB 1K88 - caspase 7 / PDB 1F1J - caspase 9 / PDB 1NW9 |
| 8. Liens Internet et références bibliographiques |
|
Keilin, D. (1925) Proc. R. Soc. (Lond.) B. Biol. Sci 98, 312-339 Ambler, R.P. (1991). "Sequence variability in bacterial cytochromes c" Biochim. Biophys. Acta 1058, 42 - 47 |
|
|
"Nomenclature of Electron-Transfer Proteins : section 4 - Cytochromes" "Reproductive and Cardiovascular Disease Research Group" |
|
|
Pokkuluri et al. (2011) "Structure of a novel dodecaheme cytochrome c from Geobacter sulfurreducens reveals an extended 12 nm protein with interacting hemes" J. Struct. Biol. 174, 223 - 233 Allen et al. (2008) "Order within a mosaic distribution of mitochondrial c-type cytochrome biogenesis systems ?" FEBS J. 275, 2385 - 2402 |
|
|
Lynes & Simmen (2011) "Urban planning of the endoplasmic reticulum (ER): How diverse mechanisms segregate the many functions of the ER" BBA 1813, 1893 - 1905 Rautureau et al. (2010) "Intrinsically Disordered Proteins in Bcl-2 Regulated Apoptosis" Int. J. Mol. Sci. 11, 1808 - 1824 |
|
|
"Les mécanismes moléculaires de l'apoptose" - J.E. Ricci "The CAspase Substrate dataBAse Homepage" "CASVM: Server for SVM Prediction of Caspase Substrates Cleavage Sites" "Domain structure of the human caspases in the UniProt database" Denault & Salvesen (2002) "Caspases" Curr. Protoc. Prot. Sci. 21.8 |
|
![]()