| La dégradation des protéines : marquage par l'ubiquitine et les protéasomes |
| Tweet |
|
|
1. Généralités 2. Le marquage des protéines à dégrader par l'ubiquitine
3. Les enzymes de désubiquitination |
4. Le protéasome 26S 5. Le protéasome 20S 6. Le protéasome 19S 7. Autres activateurs du protéasome 20S 8. Les machineries protéolytiques de la famille [AAA / ATPase] 9. La protéase ATP-dépendante bactérienne ClpXP 10. Liens Internet et références bibliographiques |
1. Généralités. Les protéines sont constamment synthétisées et dégradées dans tous les types de cellules dans le but :
Il existe 3 processus de protéolyse (l'hydrolyse des protéines). a. Une hydrolyse non spécifique des protéines ou des peptides alimentaires en fragments peptidiques par des protéases (enzymes digestives). Il en existe plusieurs familles constituées d'un trés grand nombre d'enzymes : |
|
aspartyl-protéases |
protéases à thréonine |
|||
et bien d'autres ... |
et bien d'autres ... |
et bien d'autres ...
|
et bien d'autres ... |
et bien d'autres ... |
Toutes les informations fonctionnelles (spécificité des substrats, inhibiteurs, séquences et autres) et structurales sont recensées dans les bases de données, par exemple : The MEROPS database. |
b. Un système protéolytique dans le lysosome des Eucaryotes qui permet de recycler les protéines membranaires, les protéines extracellulaires et les protéines caractérisées par de trés longs temps de demi-vie. c. Enfin, chez les Eucaryotes, les Archées et certaines bactéries, il existe un processus hautement sélectif et régulé qui se déroule dans le noyau et le cytoplasme et qui nécessite de l'énergie (hydrolyse de l'ATP) : il s'agit du système protéolytique ubiquitine-protéasome. Ce système joue un rôle majeur dans la dégradation des protéines impliquées dans le cycle cellulaire, la prolifération, l'apoptose, le contrôle de la transcription, la transduction du signal et la régulation du métabolisme. De nombreuses enzymes qui constituent des points de contrôle des voies métaboliques sont aussi des cibles d'une dégradation fréquente. |
Il existe d'autres processus de protéolyse qui n'ont pas trait à la dégradation des protéines mais bien au contraire à leur maturation par modifications post-traductionnelles. Il s'agit du clivage, entre autre,
Les temps de demi-vie dans la cellule des protéines sont trés variables :
|
| Règle du N-terminal (Alexander Varshavsky, 1996) : l'acide aminé en position N-terminale a une grande importance sur le temps de demi-vie des protéines. | |
| Acide aminé N-terminal | temps de demi-vie |
| Acide aminé stabilisants | |
| Met / Gly / Ala / Ser / Thr / Val | > 20h |
| Acide aminé déstabilisants | |
| Ile / Gln | 30 min |
| Tyr / Glu | 10 min |
| Pro | 7 min |
| Leu / Phe / Asp / Lys | 3 min |
| Arg | 2 min |
| Les protéines qui possèdent un motif dit "PEST", motif de 12 acides aminés riche en Pro (P), Glu (E), Ser (S) et Thr (T), sont plus rapidement degradées. | |
Ces règles ne sont pas absolues car de nombreux paramètres influent sur le temps de demi-vie (et le taux de synthèse - dégradation) des protéines et des macromolécules biologiques. |
|
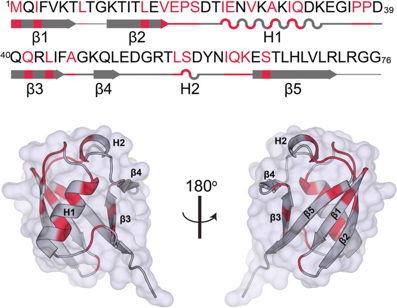
2. Le marquage des protéines par l'ubiquitine : l'ubiquitination (ou ubiquitinylation) Chez les Eucaryotes, le processus spécialisé de dégradation des protéines met en jeu l'ubiquitine, protéine de 76 acides aminés (8,6 kDa).
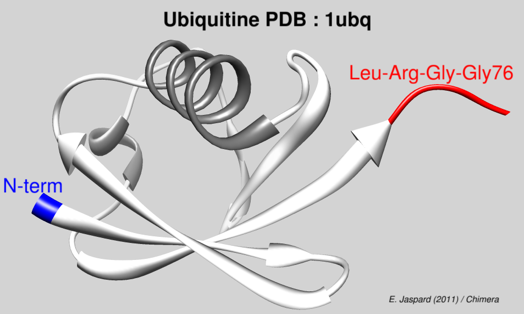
Source : Narasimhan et al. (2019) Cette protéine est ainsi appelée du fait de son ubiquité chez tous les Eucaryotes et dans tous les compartiments cellulaires des Eucaryotes. Le mécanisme complet dans lequel cette protéine est impliquée a été élucidé au début des années 80 par Aaron Ciechanover, Avram Hershko et Irwin Rose (Prix Nobel de Chimie en 2004). L'ubiquitine mature contient en particulier 7 résidus lysine et la glycine 76 C-terminale qui jouent un rôle trés important : MQIFVK6TLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIFAGKQLEDGRTLSDYNIQK63ESTLHLVLRLRGG76 L'ubiquitine (code PDB : 1UBQ) est toujours synthétisée sous la forme d'un précurseur inactif avec une extension du côté C-terminal au delà de la glycine 76.
|
|
Visualisation de l'ubiquitine de Homo sapiens à une résolution de 1,8 Å (code 1UBQ).
|
Il existe des protéines appelées "Ubiquitin-like protein" (UBLs) qui marquent également des protéines impliquées dans diverses voies de transduction du signal :
|
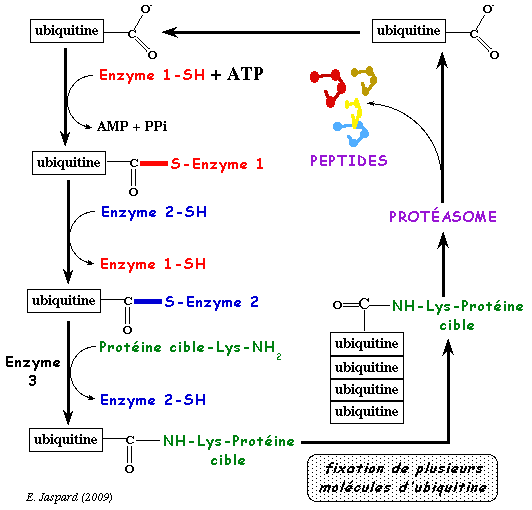
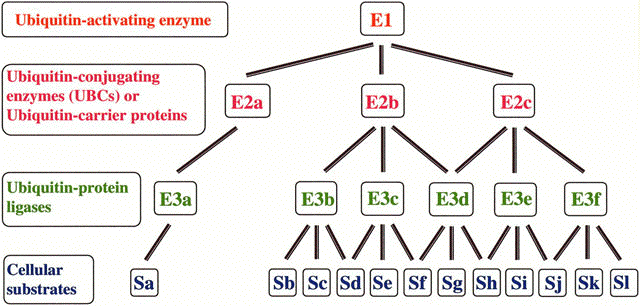
b. Mécanisme du marquage par l'ubiquitine Les protéines marquées sont attachées covalamment à un grand nombre de molécules d'ubiquitine par un système multi-enzymatique qui inclut :
Le mécanisme du marquage de la séquence d'une protéine par l'ubiquitine ou ubiquitination est présenté dans la figure ci-dessous :
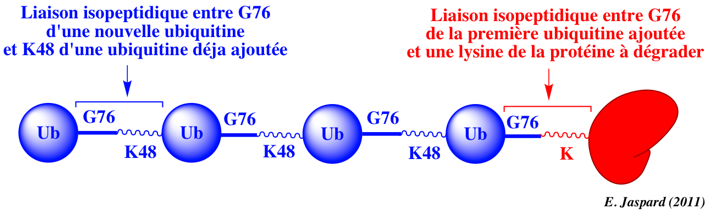
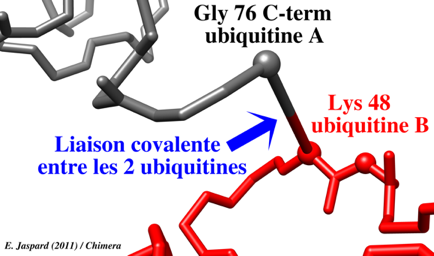
a. Le groupement carboxyle libre de la glycine 76 C-terminale (G76) d'une molécule d'ubiquitine est activé par la cystéine (-SH) du site actif de l'enzyme E1, sous la forme d'une liaison thioester à haut potentiel énergétique. Cette étape requiert de l'énergie (hydrolyse de l'ATP). b. Cette liaison thioester est ensuite transferée à une cystéine réactive de l'enzyme E2. c. Finalement, le groupement carboxyle de G76 de l'ubiquitine est lié via une liaison amide (isopeptidique) au groupement ε-aminé de la chaîne latérale d'un résidu lysine (K) de la protéine cible à dégrader. Cette étape est catalysée par l'enzyme E3. d. Si la protéine est déstinée à être dégradée, ce processus est répété. En effet, il faut au moins 4 molécules d'ubiquitine avant que la protéine à dégrader soit dirigée vers le protéasome 26S pour y être trés rapidement hydrolysée en peptides de 3 à 24 acides aminés. e. L'ubiquitine est relarguée avant l'hydrolyse et elle est recyclée. Exemple de poly-ubiquitination 1 molécule d'ubiquitine est ajoutée à la protéine à dégrader via une liaison isopeptidique entre G76 de l'ubiquitine et le groupement ε-aminé de la chaîne latérale d'un résidu lysine de la protéine à dégrader (figure ci-dessous).
Puis 3 molécules d'ubiquitine sont ajoutées via le même type de liaison entre G76 de l'ubiquitine nouvellement ajoutée et K48 de la molécule d'ubiquitine précédemment ajoutée. Il peut cependant se former différents types de polymères de topologies variées (voir le devenir des chaînes marquées par l'ubiquitine). |
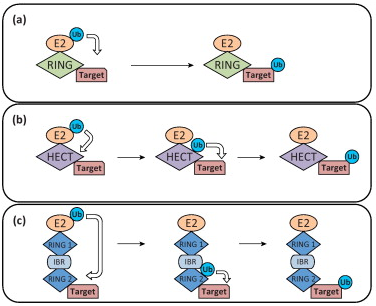
d. Les enzymes E3 - "HECT-type E3 ubiquitin transferase" Il existe 3 principaux groupes d'enzymes E3 chez les eukaryotes. α. Le groupe qui contient un domaine structural appelé RING ("Really Interesting New Gene").
Source : Deshaies & Joazeiro (2009)
β. Le groupe qui contient un domaine structural appelé HECT ("Homologous to the E6-AP Carboxyl Terminus"). γ. Le groupe qui a une architecture "RING-between-RING" (RBR). Ces enzymes E3 possèdent 2 domaines RING : l'un interagit avec une enzyme de conjuguaison E2 et l'autre possède l'activité ligase comme les enzymes E3 à domaine HECT.
Source : Rieser et al. (2013) |
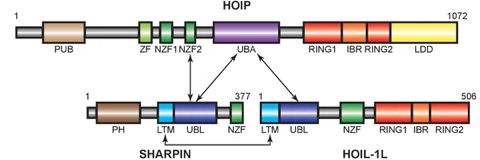
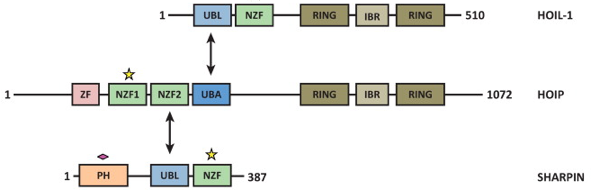
e. Illustration : le complexe LUBAC LUBAC ("Linear Ubiquitin Chain Assembly Complex") est un complexe protéique à activité ubiquitine ligase E3 à domaine "RING-between-RING" qui synthètise des chaînes linéaires de poly-ubiquitines. Le complexe LUBAC est un énorme complexe d'environ 600 kDa (figure ci-dessous).
Source : Carvajal et al. (2021) Le complexe LUBAC est constitué de 3 protéines :
Le premier domaine NZF1 ("Nuclear protein localisation 4 (Npl4)-Zinc-Finger 1") de HOIP et chacun des domaines NZF de SHARPIN et HOIL-1 sont impliqués dans la fixation à l'ubiquitine. Les autres domaines sont impliqués dans la formation du complexe LUBAC. HOIP (partie centrale de LUBAC) se fixe à HOIL-1 et à SHARPIN. L'interaction [HOIP/HOIL-1] et [HOIP/SHARPIN] s'effectue via un domaine structural qui mime la structure de l'ubiquitine : le domaine "UBiquitin-Like" (UBL).
Source : Rieser et al. (2013) - UBA : "UBiquitin-Associated domain" L'assemblage des chaînes linéaires de polyubiquitines par LUBAC a 2 caractéristiques :
|
|
f. Devenir des protéines selon le type de marquage par l'ubiquitine (mono- et poly-ubiquitination greffée ou linéaire) Les 7 lysines de l'ubiquitine peuvent être utilisées pour le marquage. Cette multiplicité de topologies des chaînes explique en partie la diversité de signaux médiées par le marquage par l'ubiquitine : certaines protéines sont destinées à la dégradation par le protéasome et d'autres protéines sont destinées à des voies de signalisation. Le devenir du complexe [protéine marquée - molécules d'ubiquitine] dépend notamment :
Protéines déstinées à la dégradation Les chaînes constituées d'au moins 4 molécules d'ubiquitine où G76 d'une molécule d'ubiquitine est attaché à K48 de la molécule d'ubiquitine suivante (voir schéma plus haut) sont dirigées vers le protéasome 26S pour y être dégradées.
|
Protéines déstinées à des voies de signalisation α. Les protéines mono-ubiquitinylées (fixation d'une molécule d'ubiquitine sur un résidu de la protéine à dégrader) ou multi-mono-ubiquitinylées (fixation d'une molécule d'ubiquitine sur plusieurs résidus de la protéine à dégrader) ne sont pas destinées à la dégradation par le protéasome 26S. Exemples :
β. Les protéines sur lesquelles s'est fixée une courte chaîne de molécules d'ubiquitine liées entre elles par K6 ou par K63 ne sont pas destinées à la dégradation par le protéasome 26S. Exemples :
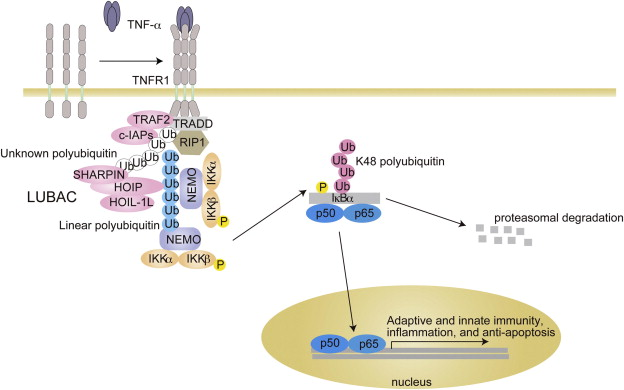
γ. Illustration de 2 voies d'activation du facteur NF-κB ("Nuclear Factor-κB") NF-κB est un facteur de transcription activé par de nombreux stimuli tels que les infections virales ou fongiques, les agents pathogènes bactériens, les cytokines proinflammatoires, les agents génotoxiques, les rayons ultraviolet ou le stress oxidatif. La voie classique (figure ci-dessous) est activée par des cytokines inflammatoires comme TNF-α : le résultat est l'activation du complexe IKK (IKKα, IKKβ et NEMO). Le complexe IKK phosphoryle IκBα, entraînant son marquage par l'ubiquitine et sa dégradation par le protéasome : NF-κB est ainsi libéré. NF-κB est constitué de p50 et p65 qui sont transloqués dans le noyau où ils activent la transcription de nombreux gènes.
Source : Tokunaga & Iwai (2012) La voie non-classique implique l'activation du dimère IKKα qui phosphoryle p100. p100 est alors maturé en p52 par la voie [ubiquitine / protéasome]. Le facteur NF-κB activé par cette voie non-classique est constitué de p52 et RelB qui sont également transloqués dans le noyau. |
|
3. Les enzymes de désubiquitination Il existe des dizaines d'enzymes qui catalysent la désubiquitination ("DeUBiquitinating enzymes" - DUB). On recence au moins six groupes de DUB : Ubiquitin C-terminal hydrolases (UCH); Ubiquitin-specific processing proteases (UBP or USP); Jab1/Pad1/MPN-domain-containing metallo enzymes (JAMM); Otu-domain ubiquitin-aldehyde-binding proteins (exemple : Ubiquitin thioesterase OTUB1); Ataxin-3/Josephin; ubiquitin-like proteases (ULP). Ce sont d'importants régulateurs du système ubiquitine - protéasome. Ces enzymes :
|
|
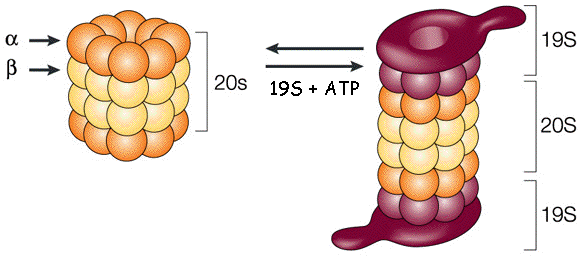
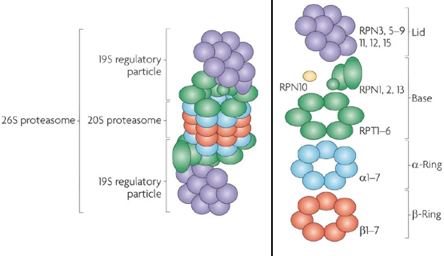
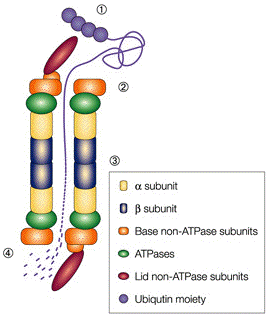
Les protéasomes sont des complexes protéiques que l'on trouve chez tous les Eucaryotes et les Archées et chez certaines bactéries. Chez les Eucaryotes, les protéasomes sont situés dans le noyau et dans le cytoplasme. Le protéasome 26S est la forme la plus fréquente du protéasome. Le protéasome 26S est un énorme complexe multi-enzymatique constitué de nombreuses sous-unités : il résulte de l'assemblage du protéasome 20S et du protéasome 19S qui met en jeu de l'énergie via l'hydrolyse de l'ATP.
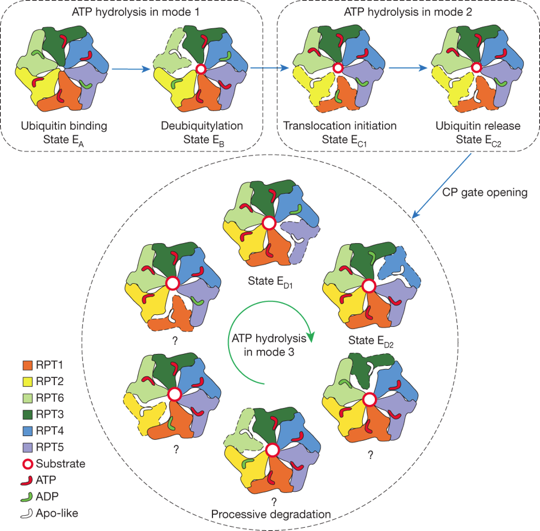
Source : Julian Adams (2004) Le protéasome 26S est tellement volumineux (environ 3000 kDa) qu'il s'apparente davantage à une particule qu'à une molécule. C'est la raison pour laquelle on le désigne par le symbole "S" ou Svedberg, l'unité du coefficient de sédimentation. Ci-dessous, schéma des 3 principaux modes d'hydrolyse coordonnée de l'ATP et description de la manière dont ils régulent le cycle complet du traitement du substrat par la forme holoenzyme du protéasome.
Source : Dong et al. (2019) L'holoenzyme est assemblée à partir :
La reconnaissance des substrats ubiquitinylés est assurée par les récepteurs de l'ubiquitine RPN1, RPN10 et RPN13. Lorsqu'une protéine substrat est fixée au RP, les domaines globulaires de cette protéine substrat sont dépliés par un "moteur ATP" hétérohexamérique :
|
|
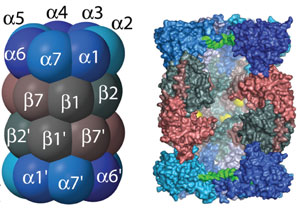
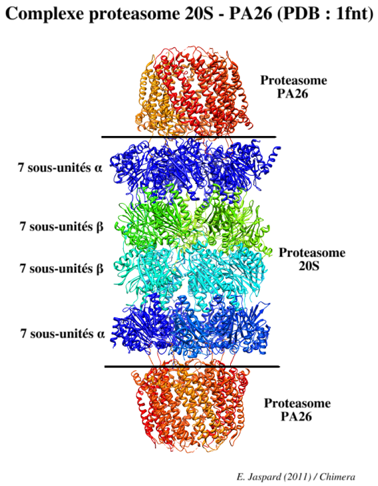
Le protéasome 20S est le coeur protéolytique du protéasome 26S (figure ci-dessous).
Source : Groll et al. (1997) Ces sites actifs sont donc confinés à l'intérieur du protéasome 20S et isolés de l'environnement cellulaire. Ainsi, les protéines de la cellule qui ne doivent pas être dégradées sont protégées d'une dégradation inopportune. L'accès des protéines à dégrader au site actif de ce complexe est sous le contrôle des sous-unités α qui ne permettent l'accès qu'aux polypeptides qui ont été préalablement dépliés (voir le protéasome 19S). Les sous-unités β1, β2 et β5 possèdent l'activité protéolytique et elles hydrolysent la liaison peptidique de manières spécifiques :
Les sous-unités β1, β2 et β5 sont synthétisées sous forme de précurseurs activés après protéolyse d'un peptide N-terminal. Ces hydrolases constituent une famille unique de protéase à thréonine. Les sous-unités β3, β4, β6 et β7 n'ont pas d'activité protéolytique. |
|
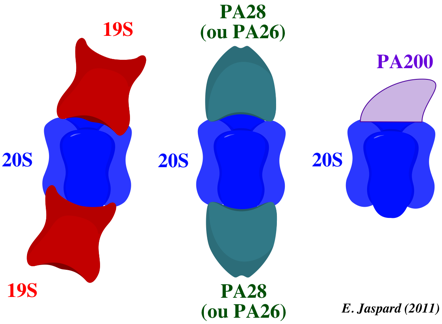
Le protéasome 19S est aussi appelé PA700 ("Proteasome Activator MW 700") ou "regulatory complex". Il forme une coiffe à une ou aux deux extrémité(s) du protéasome 20S.
Source : Murata et al. (2009)
|
|
Les rôles du protéasome 19S sont :
Source : Sullivan et al. (2003) 1. La reconnaissance et la fixation de la protéine qui doit être dégradée. 2. L'élimination des molécules d'ubiquitine (qui sont recyclées) par les sous-unités isopeptidases. Il existe deux classes d'isopeptidases : les "Ubiquitin C-terminal Hydrolases" (UCH) et les "UBiquitin-specific Proteases" (UBP). 3. Le dépliement de la chaîne polypeptidique de la protéine qui doit être dégradée car le diamètre (1 nm) du "conduit d'entrée" au protéasome 20S et le diamètre de la cavité interne (5 nm) ne permettent pas au protéasome 20S d'encapsuler des protéines repliées : les 6 sous-unités ATPases du protéasome 19S catalysent le dépliement grâce à l'énergie produite par l'hydrolyse de l'ATP. 4. L'activation du protéasome : ces ATPases sont également responsables de l'ouverture de la "barrière" qui ferme l'entrée du protéasome 20S et de l'injection des protéines dépliées dans la cavité protéolytique. Pour ce dernier rôle, le protéasome 19S est aussi appelé "Proteasome Activator" (ou PA700). |
8. Les machineries protéolytiques de la famille [AAA / ATPase] La famille AAA maintenant appelée AAA+ (ATPases Associées à diverses Activités cellulaires - "ATPases Associated with diverse cellular Activities" - Kunau et al., 1993) est un groupe important d'ATPases présentes dans tous les règnes du vivant.
Il y a six clades majeurs dans la famille des protéines à domaines AAA+ : les sous-unités du protéasome, des métalloprotéases, les domaines D1 et D2 des ATPases à 2 domaines AAA, le groupe [MSP1 / katanine / spastine] et BCS1 et ses homologues. Exemples de rôles biologiques des protéines [AAA / ATPases]
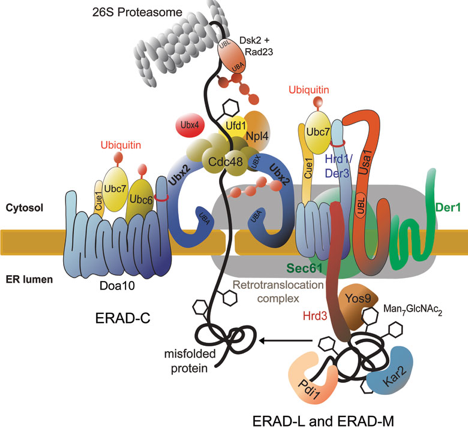
Les protéines sécrétées mal repliées sont exportées depuis le réticulum endoplasmique (RE) puis dégradées par la voie de dégradation associée au RE (voie ERAD).
Source : Eisele et al. (2010) Les protéines membranaires ou luminales non fonctionnelles sont extraites du RE puis dégradées dans le cytosol par les protéasomes. L'extraction et la rétro-translocation de ces protéines est assistée par le complexe [Cdc48p(Ufd1p/Npl4p)] du côté cytosolique de la membrane. Une fois dans le cytosol, ces protéines sont ubiquitinylées avant dégradation par le protéasome 26S. |
|
Les machineries protéolytiques de la famille AAA+ (ClpXP, ClpAP, ClpCP, HslUV, Lon, FtsH, PAN/20 et le protéasome 26S) assurent un contrôle qualité des protéines et sont des circuits de régulation de toutes les cellules.
|
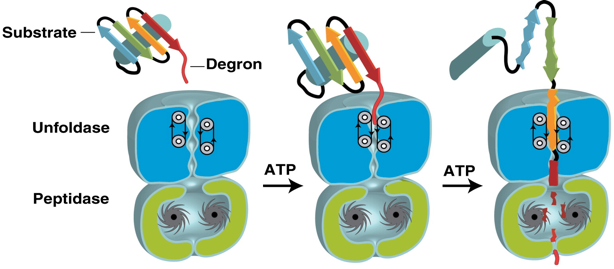
Mécanisme de base d'une protéine [AAA / ATPase] Les machineries protéolytiques de la famille AAA+ sont constituées :
Source : Sauer & Baker (2011)
|
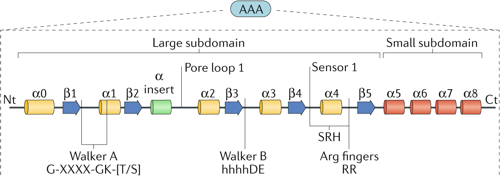
Structures quaternaires des protéines [AAA/ATPases] Les ATPases de la famille AAA+ sont caractérisées par 1 ou 2 domaine(s) conservé(s) de fixation de l'ATP d'un type appelé motif AAA. Le domaine AAA+ contient 2 sous-domaines :
Figure ci-dessous: éléments de structure secondaire conservés. Motif Walker A ([AG]-x(4)-G-K-[ST]), boucle de pore 1, motif Walker B (hhhhDE avec h = acide aminé hydrophobe, D est important pour la coordination du Mg2+ et E sert de base catalytique), capteur 1 ("sensor 1") et les doigts Arg (RR) dans la deuxième région d'homologie.
Source : Puchades et al. (2019)
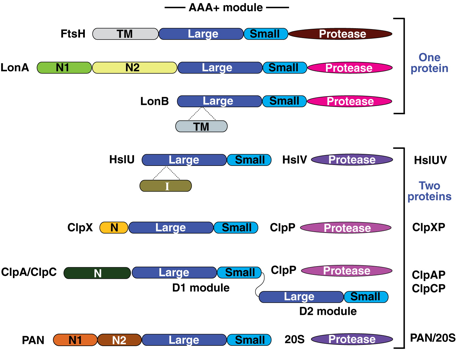
Figure ci-dessous : structures des domaines des sous-unités qui forment les sous-familles de protéases FtsH, Lon (sous-familles de LonA et LonB), HslUV, ClpXP, ClpAP, ClpCP et [PAN ("Proteasome-Activating Nucleotidase") / 20S].
Source : Sauer & Baker (2011)
|
|
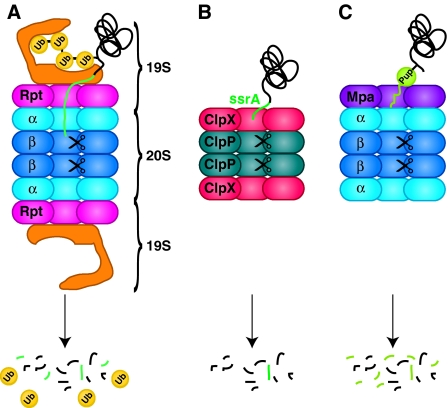
9. La protéase ATP-dépendante bactérienne ClpXP Le protéasome 26S est la seule protéase ATP-dépendante soluble dans le cytosol chez les Eucaryotes. En revanche, les bactéries possèdent plusieurs protéases ATP-dépendantes comme ClpXP, HslUV et les eubactéries possèdent le système [protéasome - ARC / Mpa] ("AAA ATPase forming Ring-shaped Complexes / Mycobacterial proteasomal ATPase"). Figure ci-dessous : comparaison de différentes classes de protéases ATP-dépendantes.
Source : Kraut & Matouschek (2010) Mpa ("Proteasome-associated ATPase") se fixe à la chaîne polypeptidique substrat via une modification covalente : la pupylation ("PUP" en vert clair).
La protéase ClpXP de Escherichia coli dégrade les protéines portant l'étiquette ssrA qui possède la séquence AANDENYALAA-COOH :
Source : McGinness et al. (2006) |
| 10. Liens Internet et références bibliographiques |
|
"Ubiquitin : molecule of the month" - Protein Data Bank Excellent site de ressources pédagogiques - Université de Bordeaux : "Ribosomes et protéasomes : la synthèse et la dégradation des protéines" Cours en ligne : "La voie de dégradation ubiquitine dépendante" - Université Jussieu |
|
|
Varshavsky A (1997). "The N-end rule pathway of protein degradation". Genes to Cells 2 (1): 13–28. Aaron Ciechanover (1998) "The ubiquitin - proteasome pathway: on protein death and cell life" The EMBO Journal 17, 7151 - 7160 Amerik A.Y. & Mark Hochstrasser, M. (2004) "Mechanism and function of deubiquitinating enzymes" BBA - Molecular Cell Research 1695, 189 - 207 |
|
|
Sullivan et al. (2003) "The diverse roles of ubiquitin and the 26S proteasome in the life of plants" Nature Reviews Genetics 4, 948 - 958 Lee & Schindelin (2008) "Structural insights into E1-catalyzed ubiquitin activation and transfer to conjugating enzymes" Cell 134, 268 - 278 |
|
|
LUBAC Kirisako et al. (2006) "A ubiquitin ligase complex assembles linear polyubiquitin chains" EMBO J. 25, 4877 - 4887 Deshaies & Joazeiro (2009) "RING Domain E3 Ubiquitin Ligases" Annu. Rev. Biochem. 78, 399 - 434 Walczak et al. (2012) "Generation and physiological roles of linear ubiquitin chains" BMC Biology 2012, 10:23 Komander & Rape (2012) "The Ubiquitin Code" Annu. Rev. Biochem. 81, 203 - 229 Tokunaga & Iwai (2012) "LUBAC, a novel ubiquitin ligase for linear ubiquitination, is crucial for inflammation and immune responses" Microbes Infect. 14, 563 - 572 Rieser et al. (2013) "Linear ubiquitination: a newly discovered regulator of cell signalling" Trends Biochem. Sci. 38, 94 - 102 |
|
|
Famille [AAA / ATPases] Kunau et al. (1993) "Two complementary approaches to study peroxisome biogenesis in Saccharomyces cerevisiae: forward and reversed genetics" Biochimie 75 , 209 - 224 Walker et al. (1982) "Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold" EMBO J. 1, 945 - 951 Sauer & Baker (2011) "AAA+ Proteases: ATP-Fueled Machines of Protein Destruction" Ann. Rev. Biochem. 80, 587 - 612 Roberts et al. (2013) "Functions and mechanics of dynein motor proteins" Nat. Rev. Mol. Cell Biol. 14, 713 - 726 Puchades et al. (2019) "The molecular principles governing the activity and functional diversity of AAA+ proteins" Nat. Rev. Mol. Cell Biol. doi: 10.1038/s41580-019-0183-6 |
|
|
Protéase bactérienne ClpXP McGinness et al. (2006) "Engineering controllable protein degradation" Mol. Cell. 22, 701 - 707 Kraut & Matouschek (2010) "Pup grows up: in vitro characterization of the degradation of pupylated proteins" EMBO J. 29, 1163 - 1164 Eisele et al. (2010) "Ubiquitylation in the ERAD Pathway" Subcell. Biochem. 54, 136 - 148 |
|
|
Groll et al. (1997) "Structure of 20S proteasome from yeast at 2.4Å resolution" Nature 386, 463-471 Julian Adams (2004) "The proteasome: a suitable antineoplastic target" Nature Reviews Cancer 4, 349 - 360 Murata et al. (2009) "Molecular mechanisms of proteasome assembly" Nat. Rev. Molec. Cell Biol. 10, 104 - 115 Förster et al. (2010) "Towards an integrated structural model of the 26S proteasome" Mol. Cell. Proteomics 9, 1666 - 1677 |
|
|
Dong (2019) "Cryo-EM structures and dynamics of substrate-engaged human 26S proteasome" Nature 565, 49 - 55 Narasimhan et al. (2019) "DNP-Supported Solid-State NMR Spectroscopy of Proteins Inside Mammalian Cells" Angew. Chem. Int. Ed. Engl. 58, 12969 - 12973 Carvajal et al. (2021) "The linear ubiquitin chain assembly complex (LUBAC) generates heterotypic ubiquitin chains" Elife 10, e60660 |
![]()