| Dynamique de la mitochondrie : mouvement sur les microtubules, fusion et fission |
| Tweet |
|
|
1. Introduction 2. Kinesine, dyneine et dynactine : protéines motrices du transport des mitochondries 3. Les protéines clé de la fission et de la fusion des mitochondries 4. Rôle de la protéine kinase activée par l'AMP (AMPK) 5. Mouvement des mitochondrie sur les microtubules |
6. Les mitofusines MFN1 et MFN2 7. La membrane mitochondriale externe associée à la membrane du réticulum endoplasmique (MAM - "Mitochondria-Associated ER-Membrane") 8. Les sites de contacts [mitochondrie - RE] - Le complexe ERMES 9. Résumé de quelques interactions protéine-protéine 10. L'autophagie 11. Liens Internet et références bibliographiques |
|
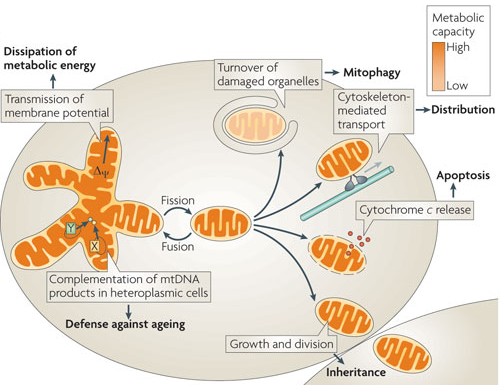
1. Introduction Les mitochondries se déplacent le long du cytosquelette jusqu'aux sites de forte demande en énergie. Leur morphologie change en réponse aux modifications de l'environnement et aux exigences de la différenciation cellulaire. Les mitochondries forment donc des réseaux très dynamiques et constituent des structures tubulaires et ramifiées. Des modèles sophistiqués ont été développés pour essayer de décrire cette dynamique. Tout au long de la vie d'un neurone, les mitochondries âgées et endommagées subissent un recyclage dynamique par fusion - fission ou par élimination par le processus appelé mitophagie (une autophagie qui dégrade les mitochondries dans les lysosomes). Le temps de demi-vie des mitochondries dans le neurone est estimé à 30 jours. Répartition de l'ADN mitochondrial
Source : Westermann B. (2010) Chaque mitochondrie contient une ou plusieurs molécule(s) d'ADN mitochondrial (ADNmt) organisée(s) en complexes [ADNmt / protéines] appelés nucléoïdes. Dans les cellules qui prolifèrent, la répartition des mitochondries dans les cellules filles doit se traduire par une bonne répartition de l'ADNmt. Chez les mammifères, les mitochondries sont transmises aux générations suivantes exclusivement par la lignée maternelle. En raison de ce mode de transmission uniparentale, l'ADNmt ne possède pas les avantages de la recombinaison résultant de la reproduction sexuée. |
Mouvements des mitochondries Protéines motrices des mitochondries :
Exemple : dans le neurone, beaucoup de mitochondries sont nécessaires près des synapses et des cônes de croissance (régions de bourgeonnement de la membrane qui forment de nouveaux axones et des dendrites) dans un neurone. Les mitochondries fusionnent ou se divisent (fission) en permanence, tant à l'intérieur des structures tubulaires qu'aux points de branchement :
Source : "Dynamic Relationship of Mitochondria and Neurons"
Une fine balance entre ces processus opposés maintient la forme appropriée, la taille et le nombre des mitochondries. Ces deux processus sont influencés par les lipides, les charges négatives et la courbure de la bicouche lipidique. Les cycles de fusion et de fission des membranes externe et interne sont nécessaires pour l'échange de l'ADN mitochondrial, des protéines et des lipides entre les mitochondries endommagées et les mitochondries saines afin de maintenir un ensemble robuste de mitochondries fonctionnelles. |
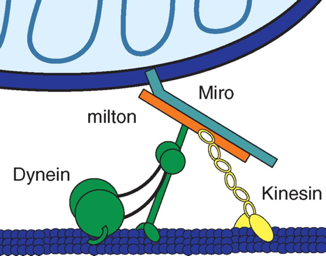
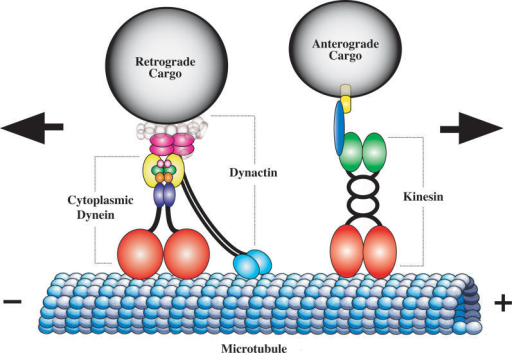
2. Kinesine, dyneine et dynactine : protéines motrices du transport des mitochondries Les kinesines ("anterograde motor kinesin-1" ou "kinesin heavy chain" - Kif5) :
Les dyneines (associées à la dynactine - "retrograde motor [dynein/dynactin] complex") :
Source : Schwarz TL (2013)
La dynactine est un complexe protéique multimérique composé notamment de :
Source : Cell Biology group project La kinesine 1 et le complexe [dyneine/dynactine] sont liées aux mitochondries par leurs interactions avec 2 protéines spécifiques de la mitochondrie : Miro et Milton.
|
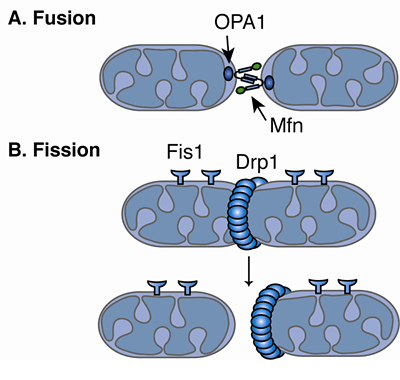
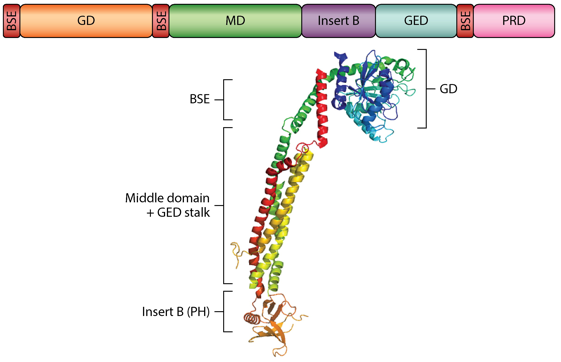
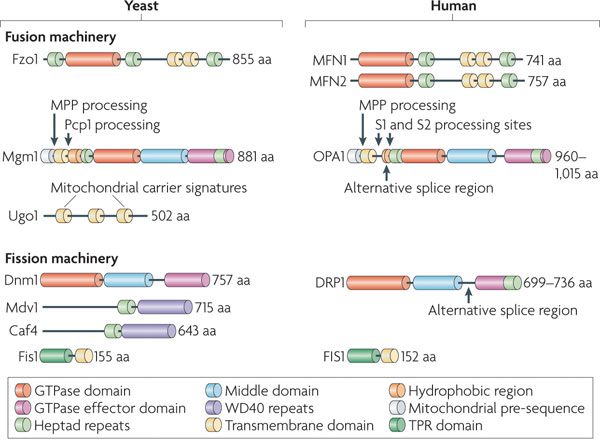
3. Les protéines clé de la fusion et de la fission des mitochondries Les dynamines forment une superfamille de protéines qui comprend : les dynamines, les protéines "dynamin-like", OPA1, les protéines Mx ("interferon-induced GTPases"), les mitofusines/Fzo1 et les atlastines. Elles sont impliquées notamment dans le bourgeonnement des vésicules de transport, la division des organites, la résistance aux pathogènes. Elles sont associées aux clathrines et participent à la formation de vésicules lors de l'endocytose. Les dynamines sont des GTPases qui s'auto-assemblent en hydrolysant le GTP : l'auto-assemblage nécessite l'hydrolyse du GTP et la formation de structures d'ordre supérieur stimule l'hydrolyse du GTP. La fusion et la fission sont régulées par des protéines apparentées à la dynamine ("Dynamin Related Proteins" - DRP). Les DRP possèdent plusieurs domaines :
Source : Labbé et al. (2014)
PRD : "Proline-Rich Domain" / PH : "Pleckstrin Homology" |
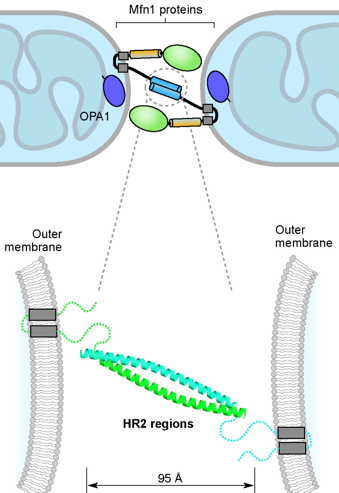
a. La fusion des mitochondries
Source : Westermann B. (2010) |
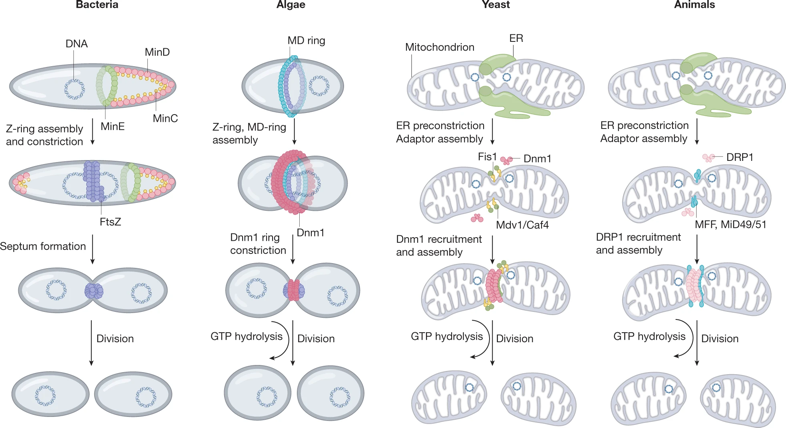
b. Les machineries de division chez certaines bactéries, algues et eucaryotes La division est un processus par étapes qui implique l'assemblage d'un anneau contractile au niveau de la membrane et la fission ultérieure. Le divisome désigne l'ensemble des entités biologiques impliquées dans la division / fission;
Source : Kraus et al. (2021) |
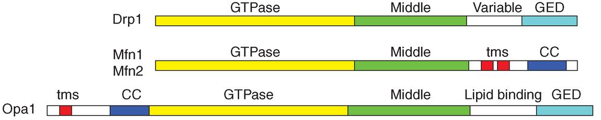
c. La fission des mitochondries
Source : van der Bliek et al. (2013)
|
| Levure | Homme - mammifères | ||
| Fusion | Fission | Fusion | Fission |
| Fzo1 (membrane externe) | Dnm1 est recrutée par Fis1 (membrane externe) | MFN1 et MFN2 (membrane externe) | Drp1 est recrutée par Fis1 |
| Mgm1 (membrane interne) | Les protéines Mdv1 et Caf4 servent d'adapteurs. | OPA1 (membrane interne) | Pas d'adapteurs identifiés |
| Fzo1 et Mgm1 sont connectées par la protéine Ugo1 (membrane externe) | --------------- | Pas d'équivalent à Ugo1 identifié | --------------- |
| La protéase membranaire Pcp1 est impliquée dans l'épissage alternatif de Mgm1 | --------------- | PARL est impliquée dans la maturation de OPA1 | --------------- |
| MPP ("Matrix-Processing Peptidase") hydrolyse la séquence d'adressage dans la matrice mitochondriale après l'import | |||
4. Rôle de la protéine kinase activée par l'AMP (AMPK) La chaîne respiratoire des mitochondries est le siège de la synthèse de l'ATP. La protéine kinase activée par l'AMP (AMPK) est un senseur métabolique capital de la variation de l'énergie d'une cellule. L'activation de l'AMPK :
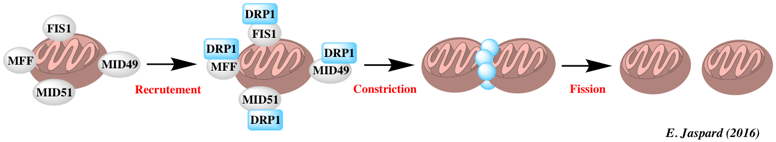
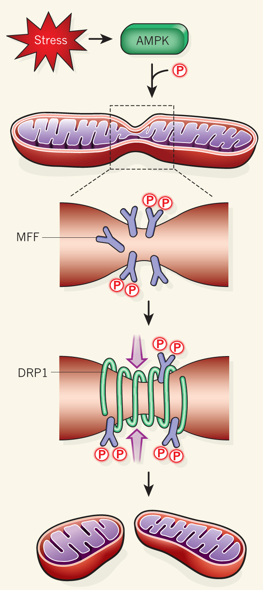
L'AMPK est rapidement activée en réponse aux drogues qui inhibent la chaîne respiratoire (la roténone et l'antimycine A). Cette activation est probablement induite par l'augmentation du rapport des concentrations [AMP/ATP]. Drp1 est le facteur mitochondrial clé de la fission en entourant les sites de constriction de la membrane mitochondriale et en formant les complexes en spirale qui médient la scission mitochondriale. DRP1 est principalement situé dans le cytoplasme. Cependant Drp1 est recruté sur les futurs sites de scission par un récepeur situé sur la membrane externe : le facteur de fission MFF ("Mitochondrial Fission Factor") qui intervient aussi dans la fission des peroxisomes. Deux autres récepteurs situés sur la membrane externe participent au recrutement de Drp1 : il s'agit de MID49 et de MID51 ("Mitochondrial Dynamics protein of 49 kDa" et "Mitochondrial Dynamics protein of 51 kDa").
L'AMPK phosphoryle S155 et S172 de MFF ce qui augmente le recrutement de Drp1. Ce mécanisme semble dont réguler la forme des mitochondries. L'augmentation de la fission induite par l'AMPK facilite l'élimination sélective des mitochondries endommagées par le processus d'autophagie (mécanisme de contrôle qualité en réponse à l'action de poisons).
Source : Wang & Youle (2016) |
5. Mouvement des mitochondrie sur les microtubules a. Les protéines Park, PINK1 et Clu Le gène PARK2 est l'un des plus longs gènes du génome de l'homme. Ce gène code pour une protéine appelée Park2 ou parkin ("E3 ubiquitin-protein ligase parkin"). La parkin est une ubiquitine ligase E3 qui joue un rôle dans la machinerie cellulaire qui dégrade les protéines non nécessaires (endommagées ou en excès) par le marquage par l'ubiquitine puis la dégradation par les protéasomes. PINK1 ("Phosphatase and TENsin homolog (PTEN)-INduced putative Kinase 1") est une [Ser/Thr] protéine kinase nécessaire au maintien de la morphologie et du fonctionnement de la mitochondrie, éventuellement en facilitant la [fission / fusion] mitochondriale.
Les études de la parkin et de PINK1 de la drosophile renforcent l'hypothèse que les disfonctionnements mitochondriaux sont au cœur de l'étiologie de la maladie de Parkinson. Protéine Clu ("Clustered mitochondria protein homolog")
|
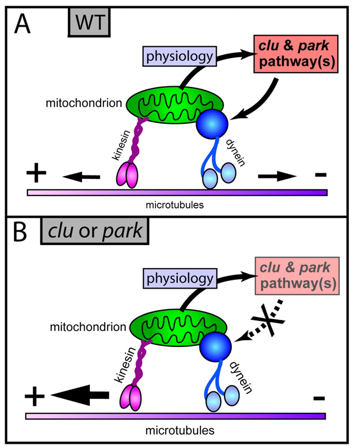
b. Modèle proposé pour le fonctionnement de la voie Clu et Park En conditions normales, la voie Clu et Park serait un senseur de l'état physiologique des mitochondries : elle adapterait en conséquence les mouvements des mitochondries le long des microtubules. Par exemple :
La voie Clu et Park semble donc impliquée dans la santé de la mitochondrie et de la cellule en contrôlant le positionnement des mitochondries sur les microtubules, le cytosquelette fournissant le moyen de distinguer les mitochondries correctes des mitochondries défectueuses.
Source : Cox & Spradling (2009) La voie Clu et Park a lieu dans le cytoplasme et a pour finalité de détecter l'état physiologique de la mitochondrie et d'adapter son mouvement sur les microtubules : le mouvement de la mitochondrie est contrôlé par les moteurs kinésine ("plus-end-directed" - en rose) et les moteurs dynéine ("minus-end-directed " - en bleu). (A) En conditions normales, la modulation des activités relatives [kinésine / dynéine] par la voie Clu et Park permet à la mitochondrie de se déplacer dans les deux sens (flèches de taille égale vers la droite et vers la gauche). (B) Quand la voie Clu et Park est défectueuse (ou que les niveaux d'espèces réactives de l'oxygène sont élevés), le mouvement vers l'extrémité négative ("minus-end-directed") des microtubules est bloquée : les mitochondries se déplacent alors préférentiellement vers l'extrémité positive des microtubules (flèche plus importante). Dans d'autres cellules, la voie Clu et Park pourrait réguler le mouvement mitochondrial par le biais d'autres types de protéines motrices. |
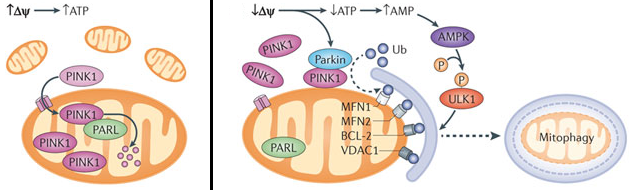
c. Interaction PINK1, parkin et mitophagie Le potentiel transmembranaire (Δψm) est utilisé pour la synthèse d'ATP et pour l'import de protéines telles que PINK1.
Source : Galluzz et al. (2012)
L'ensemble de ces événements stimulent la mitophagie. PARL ("Presenilin-Associated Rhomboid-Like") : elle est essentielle pour la protéolyse d'une forme anti-apoptotique de OPA1 qui empêche la libération de cytochrome c mitochondrial en réponse aux signaux apoptoptiques intrinsèques. Elle favorise les changements de la morphologie des mitochondries. LONP1 ("LON peptidase 1") est une protéase à serine. |
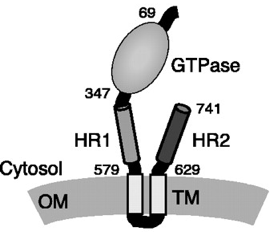
6. Les mitofusines MFN1 et MFN2 MFN1 et MFN2 :
La mitofusine 1 (MFN1 - 741 acides aminés) :
Source : Chan D.C. (2006)
Source : Koshiba et al. (2004) La mitofusine 2 (MFN2 - 757 acides aminés) contribue à déterminer la morphologie des mitochondries et à les attacher au réticulum endoplasmique (voir MAM). MFN2 est :
La protéine MITOL ("E3 ubiquitin-protein ligase MARCH 5" - (EC 6.3.2), située dans membrane externe mitochondriale, régule la dynamique mitochondriale, en interagissant avec MFN2. Elle participe à l'ubiquitinylation de Fis1, Dnm1L et MFN1. |
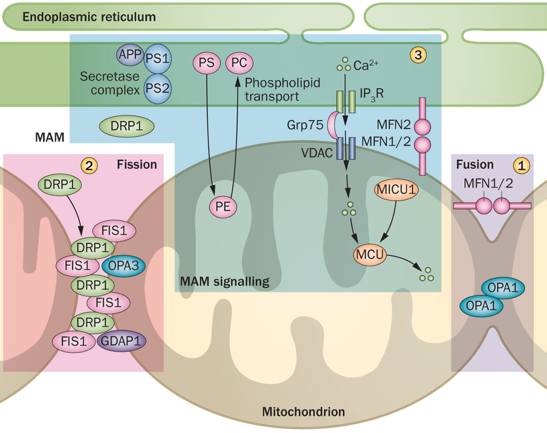
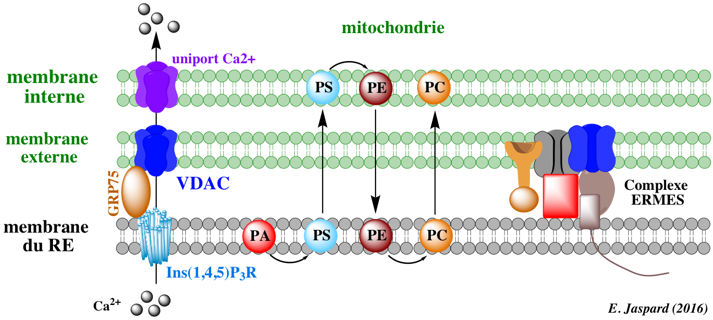
7. La membrane mitochondriale externe associée à la membrane du réticulum endoplasmique C'est un mécanisme de communication entre le réticulum endoplasmique (RE) et les mitochondries. Les sites de contact entre le RE et les mitochondries semblent nécessaires pour réguler la dynamique et la morphologie des mitochondries.
Source : Burté et al. (2015)
MFN2 est une protéine de la membrane externe des mitochondries : cependant, la MAM est enrichie environ 14 fois en MFN2. En revanche, MFN1 (qui peut former un complexe avec MFN2) n'est présente que dans la membrane des mitochondries. Abréviations de la figure ci-dessus : APP : amyloid precursor protein / GDAP1 : Ganglioside-induced differentiation-associated protein 1 / Grp75 : 75 kDa glucose-regulated protein / IP3R : inositol 1,4,5-trisphosphate receptor / MCU : mitochondrial calcium uniporter protein / MFN : mitofusin / MICU1 : mitochondrial calcium uptake protein 1 / PC : phosphatidylcholine / PE : phosphatidylethalonamine / PS : phosphatidylserine / PS1 : presenilin 1 / PS2 : presenilin 2 / VDAC : voltage-dependent anion channel / γ-sécretase : complexe de protéases qui contient entre autres la presenilin 1 / Fis1 ("mitochondrial fission 1 protein") : protéine de la membrane externe mitochondriale - composant du complexe "ARCosome" |
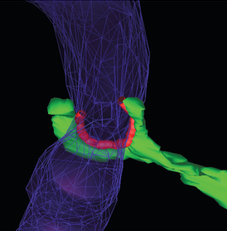
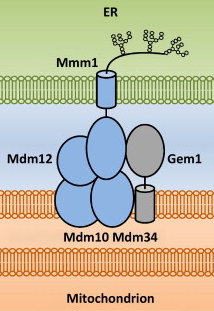
8. Les sites de contacts [mitochondrie - RE] - Le complexe ERMES Cliché ci-dessous : la technique de tomographie électronique révèle la structure tridimensionnelle des sites de contacts (en rouge) entre les mitochondries (en violet) et les tubules du RE (en vert) chez la levure.
Sources : Phillips & Voeltz (2016) - Cliché original : Friedman et al. (2011) L'ensemble des sites de contact (qui peuvent représenter de 2 à 5% de la surface d'une mitochondrie, chez les mammifères) rapproche les deux organites à une distance de 6 à 15 nm. Une si petite distance permet l'échange de lipides et de calcium via des canaux. Chez la levure Saccharomyces cerevisiae, un complexe protéique appelé ERMES ("ER-Mitochondria Encounter Structure") établit de multiples sites de contact entre le RE et les mitochondries. Chez Saccharomyces cerevisiae, ERMES est composé de 5 protéines :
Source : Klecker et al. (2014)
Chez les animaux :
|
| Nom et fonctions des protéines impliquées dans les sites de contacts [mitochondrie - RE] chez les mammifères et la levure | ||
MFN1 - MFN2 |
MFN1 : protéine de la mitochondrie |
transfert de calcium au niveau des sites de contacts [mitochondrie - RE] |
| VDAC - GRP75 - Ins(1,4,5)P3R | VDAC : protéine de la membrane externe mitochondriale - canal calcique GRP75 : protéine soluble Ins(1,4,5)P3R : protéine membranaire intégrale du RE - canal calcique |
le calcium peut être transféré du RE vers la mitochondrie : Ins(1,4,5)P3R (sortie du calcium) et VDAC (entrée du calcium) |
| PTPIP51 - VAP | PTPIP51 : protéine de la membrane externe mitochondriale avec un domaine de transfert du phosphatidylinositol phosphate VAP : protéine membranaire intégrale du RE |
transfert de lipides entre la mitochondrie et le RE |
| FIS1 - BAP31 | FIS1 : protéine de la membrane externe mitochondriale BAP31 : protéine membranaire intégrale du RE |
l'interaction FIS1 - BAP31 participe à la transmission de la mitochondrie vers le RE des signanux liés à l'apoptose |
| Mmr1 | Localisée au niveau des sites de contact du RE cortical chez la levure |
Important pour l'héritage mitochondriale chez la levure |
| complexe ERMES | Voir ci-dessus | |
| Abréviations : MFN : mitofusin / VDAC : voltage-dependent anion channel / GRP75 : glucose-regulated protein 75 / Ins(1,4,5)P3R : inositol-1,4,5-trisphosphate receptor / Mmr1 : mitochondrial Myo2p receptor-related 1 / VAP : VAMP-associated protein | ||
Les sites de contacts [mitochondrie-RE] sont riches en lipides et en enzymes de synthèse des lipides.
Le complexe ERMES contribue à ce processus. |
| 9. Résumé de quelques interactions protéine-protéine | ||||
|
Drp1 ("Dynamin related protein 1, isoform A" - EC 3.6.5.5 - Drosophila melanogaster) interagit avec : |
|
Dnm1 ("Dynamin-related protein 1" - équivalent de Drp1 chez la levure) interagit avec : |
|
|
| OPA1 ("OPtic Atrophy 1" - "Dynamin-like 120 kDa protein" - EC 3.6.5.5) interagit avec : |
|
MFN2 interagit avec : |
|
|
| Fis1 ("mitochondrial fission 1 protein") interagit avec : |
|
La parkin ou Park2 interagit avec : |
|
|
| Miro (RHOT1) interagit avec : |
|
ULK1 interagit avec : |
|
|
| PARL interagit avec : | Ugo1 interagit avec : Fzo1 / MGM1 / Dnm1 / DYN3 / YLH47 | Mdv1 interagit avec : Fis1 / Dnm1 / MDV1 / Fzo1 / DYN3 | ||
|
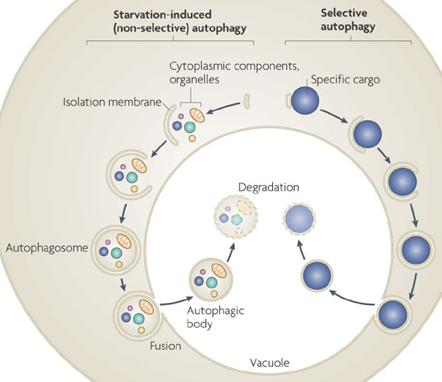
Yoshinori Ohsumi a reçu le prix Nobel de médecine en 2016 pour ses travaux sur l'autophagie. L'autophagie est un processus cellulaire qui fournit un matériel cytoplasmique au lysosome pour le recyclage. L'autophagie ou les protéines de l'autophagie interagissent avec plusieurs autres processus cellulaires tels que l'apoptose, la sécrétion et les voies de l'endocytose. L'autophagie est stimulée au-dessus du taux de base ou de repos lorsque les nutriments sont rares, lorsque les cellules sont soumis à un stress, ou quand des bactéries intracellulaires ou des organites endommagés doivent être dégradés.
Source : Nakatogawa et al. (2009) La formation de la membrane autophagique (qu'on appelle le phagophore) est initiée par 2 complexes :
Ces deux complexes sont :
|
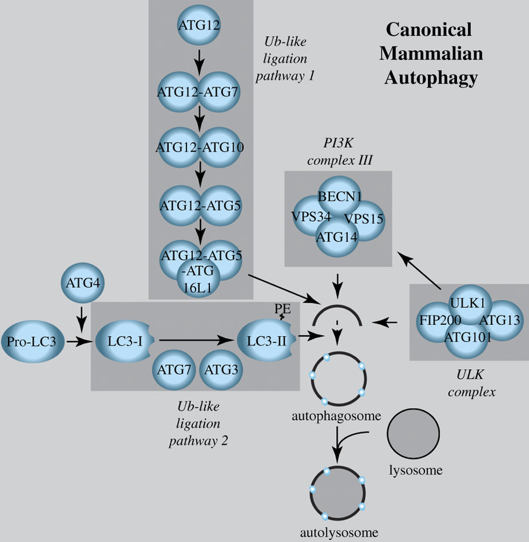
Schéma de l'autophagie chez les mammifères Les complexes ULK et PI3K III initient l'autophagie. Quand l'autophagie est stimulée, une chaîne LC3-I ("ATG4-cleaved microtubule-associated protein light chain 3") est conjuguée à la phosphatidyléthanolamine (PE) pour former LC3-II par une voie analogue à l'ubiquitine, qui comprend ATG12, ATG7, ATG10, ATG5 et ATG16L LC3-II est la forme qui permet l'allongement de la membrane du phagophore et le recrutement au phagophore.
Source : Lindqvist et al. (2015) L'autophagosome fusionne avec les lysosomes (ou la vacuole chez les plantes et la levure) pour former des autolysosomes. Ces compartiments autophagiques dégradent le matériel cellulaire enrobé. Ils renvoient des blocs de construction comme les acides aminés vers le cytoplasme.
|
| 11. Liens Internet et références bibliographiques |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
|
Koshiba et al. (2004) "Structural basis of mitochondrial tethering by mitofusin complexes" Science 305, 858 - 862 Chan DC. (2006) "Mitochondrial fusion and fission in mammals" Annu. Rev. Cell. Dev. Biol. 22, 79 - 99 Liesa et al. (2009) "Mitochondrial Dynamics in Mammalian Health and Disease" Physiol. Rev. 89, 799 - 845 Cox & Spradling (2009) "Clueless, a conserved Drosophila gene required for mitochondrial subcellular localization, interacts genetically with parkin" Dis. Model. Mech. 2, 490 - 499 Kardon & Vale (2009) "Regulators of the cytoplasmic dynein motor" Nat. Rev. Mol. Cell Biol. 10, 854 - 865 Hirokawa et al. (2009) "Kinesin superfamily motor proteins and intracellular transport" Nat. Rev. Mol. Cell. Biol. 10, 682 - 696 Nakatogawa et al. (2009) "Dynamics and diversity in autophagy mechanisms: lessons from yeast" Nat. Rev. Mol. Cell Biol. 10, 458 - 467 |
|
|
Westermann B. (2010) "Mitochondrial fusion and fission in cell life and death" Nat. Rev. Mol. Cell Biol. 11, 872 - 884 Otera et al. (2010) "Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells" J. Cell Biol. 191, 1141 - 1158 Brenner et al. (2011) "Adenine nucleotide translocase family: four isoforms for apoptosis modulation in cancer" Oncogene 30, 883 - 895 Friedman et al. (2011) "ER Tubules Mark Sites of Mitochondrial Division" Science 334, 358 - 362 Vafai & Mootha (2012) "Mitochondrial disorders as windows into an ancient organelle" Nature 491, 374 - 383 Galluzz et al. (2012) "Mitochondria: master regulators of danger signalling" Nat. Rev. Mol. Cell Biol. 13, 780 - 788 |
|
|
van der Bliek et al. (2013) "Mechanisms of mitochondrial fission and fusion" Cold Spring Harb Perspect Biol. 5, a011072 Schwarz TL. (2013) "Mitochondrial trafficking in neurons" Cold Spring Harb. Perspect. Biol. 5 Millecamps & Julien (2013) "Axonal transport deficits and neurodegenerative diseases" Nat. Rev. Neuroscience 14, 161 - 176 |
|
|
Friedman & Nunnari (2014) "Mitochondrial form and function" Nature 505, 335 - 343 Labbé et al. (2014) "Determinants and functions of mitochondrial behavior" Annu Rev Cell Dev Biol. 30, 357 - 391 Klecker et al. (2014) "Making connections: interorganelle contacts orchestrate mitochondrial behavior" Trends Cell Biol. 24, 537 - 545 Mishra & Chan (2014) "Mitochondrial dynamics and inheritance during cell division, development and disease" Nat. Rev. Mol. Cell Biol. 15, 634 - 646 |
|
|
Burté et al. (2015) "Disturbed mitochondrial dynamics and neurodegenerative disorders" Nat. Rev. Neurol. 11, 11 - 24 Lindqvist et al. (2015) "Current questions and possible controversies in autophagy" Cell Death Discov. pii: 15036 Wang & Youle (2016) "Cell biology: Form follows function for mitochondria" Nature 530, 288 - 289 Phillips & Voeltz (2016) "Structure and function of ER membrane contact sites with other organelles" Nat. Rev. Mol. Cell Biol. 17, 69 - 82 |
|
|
Mishra & Chan (2016) "Metabolic regulation of mitochondrial dynamics" J. Cell Biol. 212, 379 - 387 Cao et al. (2017) "MFN1 structures reveal nucleotide-triggered dimerization critical for mitochondrial fusion" Nature doi:10.1038/nature21077 Kraus et al. (2021) "Function and regulation of the divisome for mitochondrial fission" Nature 590, 57 - 66 |
|
![]()