
| Théorie chimio-osmotique - Synthèse d'ATP par l'ATP synthase |
| Tweet |
|
|
1. Rappel sur l'expulsion des protons au cours de la chaîne de transport d'électrons 2. La théorie chimio-osmotique et la force proton motrice 3. Le complexe V ou ATP synthase 4. Les autres types d'ATPases |
5. Expression de la force proton motrice Δp 6. Le mécanisme de la synthèse de l'ATP par l'ATP synthase 7. Transport de l'ATP formé au cours de la respiration 8. Liens Internet et références bibliographiques |
1. Rappel sur l'expulsion des protons au cours de lachaîne de transport d'électrons Voir un cours sur la chaîne repiratoire.
|
| Complexe | Protons expulsés | Remarques |
| I | 3 ou 4 protons | 3 ou 4 protons selon les données expérimentales. |
| II | pas d'expulsion | La variation d'énergie libre de la réaction est trop faible du fait d'une faible différence de potentiel de réduction entre les donneurs et les accepteurs d'électrons. |
| III | (2 x 2) protons | Variation d'énergie libre légèrement plus faible que celle générée par le complexe I. |
| IV | 2 protons | Contribue d'autant plus à la formation du gradient de concentration de protons que, du fait de la formation de molécule d'eau, il soustrait des protons de la matrice de la mitochondrie. |
| NAD(P)H déshydrogénases | pas d'expulsion | Spécifiques des plantes - Elles "court-circuitent" le premier site d'expulsion des protons (le complexe I). |
| Oxydase alternative | pas d'expulsion | Spécifique des plantes - Cette voie alternative transfère les électrons des quinones à l'oxygène et contourne ainsi 2 sites d'expulsion de protons. |
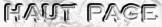
|
Dans l'enchaînement des réactions d'oxydo-réduction qui ont lieu lors du transfert des électrons de l'ensemble de la chaîne de transport d'électrons , si l'on ne considère que :
les deux demi-réaction rédox sont :
Donc : ΔG°'réaction = - n . F . ΔE°'réaction = - (2 . 96500 . 1,14) = - 220 kJ.mol-1 Puisque la variation d'énergie libre standard de la synthèse de l'ATP est environ +30,5 kJ.mol-1, la variation d'énergie libre standard liée à la réoxydation du (NADH + H+) devrait, en théorie, permettre la synthèse de : (220 kJ.mol-1 / 30,5 kJ.mol-1) = 7 molécules d'ATP par molécule de (NADH + H+) réoxydé. En fait ce sont 3 molécules d'ATP qui sont synthétisées. En effet, le réel potentiel énergétique pour la synthèse d'ATP émane de la force proton motrice. |
2. La théorie chimio-osmotique et la force proton motrice La théorie chimio-osmotique formulée par Peter Mitchell en 1961 (Prix Nobel en 1978) postule que le gradient de concentration de protons crée à travers la membrane sert de réservoir d'énergie libre pour la synthèse d'ATP. En d'autres termes :
Par rapport à l'espace intermembranaire la matrice devient :
|
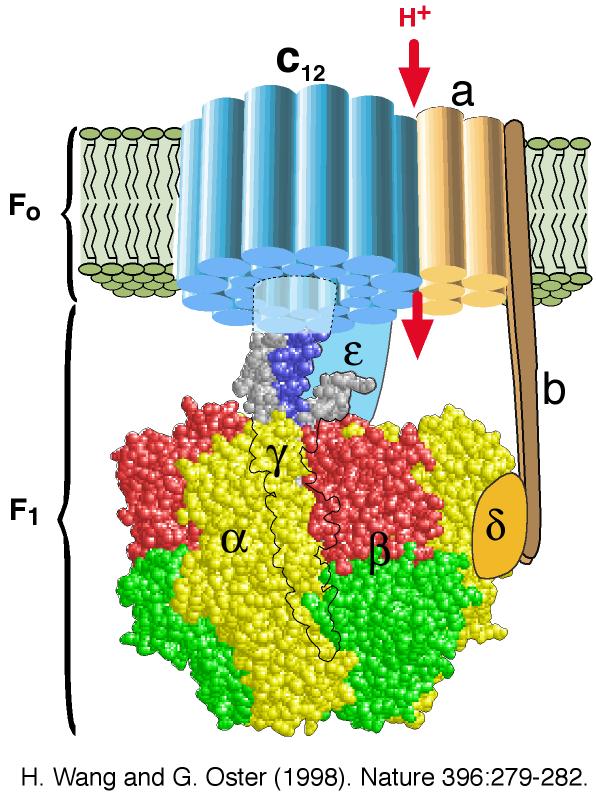
3. Le complexe V ou ATP synthase Le complexe V, appelé ATP synthase F0F1, se sert du gradient de concentration de protons comme source d'énergie pour synthétiser l'ATP : Le sigle "F" désigne un facteur de couplage, selon la nomenclature des enzymes : en effet, l'ATP synthase couple la phosphorylation de l'ADP en ATP à l'oxydation de substrats par la mitochondrie. Les ATP synthases sont des protéines oligomériques de masse molaire supérieure à 450 kDa et de structure très complexe : les types différents de chaînes polypeptidiques qui les composent sont au nombre de 8 chez E. coli, 8 ou 9 dans le chloroplaste, 10 ou plus dans la mitochondrie et au moins 13 pour celle du coeur de boeuf. F1 est l'élément qui catalyse l'hydrolyse de l'ATP : on l'appelle usuellement l'ATPase F1 ou ATP hydrolase :
F0 constitue un "tunnel" à protons sur toute l'épaisseur de la membrane interne de la mitochondrie :
|
|
Il existe plusieurs autres types d'ATPases qui n'ont pas pour fonction la synthèse d'ATP. Ce sont des ATPases ionophores ou encore appelées pompes ioniques quand l'énergie qu'elle libère en hydrolysant l'ATP est utilisée pour le transport d'ions :
|
|
A 37°C, la valeur de chaque terme est :
Remarque : certaines sources indiquent les valeurs : ΔpH = - 1 unité pH et ΔΨ = 0,15 V ou 0,16 V. La théorie chimiosmotique implique que la membrane interne est imperméable aux protons , sinon le gradient se dissiperait au fur et à mesure et effectivement elle est imperméable aux protons. |
|
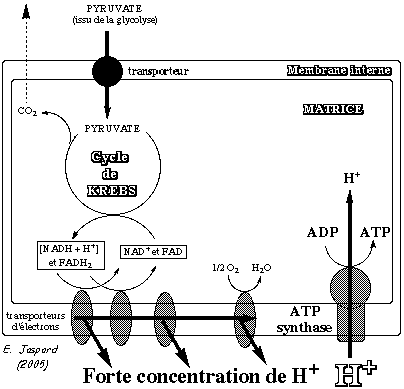
6. Le mécanisme de la synthèse de l'ATP par l'ATP synthase a. Synthèse de l'ATP L'ATP synthase est un cylindre composé de sous-unités α et β altérnées :
C'est Paul Boyer (Prix Nobel 1997) qui a proposé un modèle pour le mécanisme enzymatique de la synthèse de l'ATP (schéma ci-dessous). Ce modèle a été confirmé par l'étude de la structure de l'ATP synthase par John Walker (Prix Nobel 1997).
Dans ce schéma, les états conformationnels adoptés par les sous-unités β sont appelés :
Le mécanisme est le suivant :
Voir une animation de ce mécanisme appelé moteur moléculaire. b. Relarguage de l'ATP synthétisée au niveau de la sous-unité dans la conformation βT : Cette molécule d'ATP se trouve dans un puits thermodynamique ce qui signifie que son énergie de fixation est tellement importante qu'elle ne peut-être relarguée tant que la sous-unité est dans la conformation βT. La rotation de la sous-unité γ change la conformation des sous-unités β. Ainsi le site à haute affinité pour l'ATP (Ks < 1 µM) devient un site à faible affinité (sous-unité dans la conformation βO) et l'ATP est relargué. |
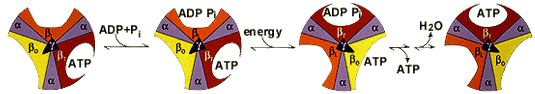
| 7. Transport de l'ATP formé au cours de la respiration Voir un cours sur les transports cellulaires. L'ATP produit dans la mitochondrie doit retourner vers le cytosol pour y être utilisé pour les divers travaux cellulaires. Inversement, l'ADP et le phosphate inorganique (Pi) issus de l'hydrolyse de l'ATP dans le cytosol, doivent entrer dans la mitochondrie pour la synthèse d'ATP.
Deux protéines de transport localisées dans la membrane interne assurent ce cycle. 1. l'adénine nucléotide translocase (transporteur ADP/ATP) est un antiport qui catalyse l'échange d'ADP pour l'ATP. Le nombre de charges négatives portées par ces 2 molécules est respectivement de 3 et 4. La force nécessaire pour cet échange [ADP3-] / [ATP4-] émane du potentiel de membrane généré au cours de la respiration (1 charge négative par molécule d'ATP). 2. le phosphate inorganique entre dans la matrice mitochondriale avec le proton (H+) via un symport neutre électriquement. La force nécessaire pour l'entrée du phosphate inorganique émane du gradient de pH (1 H+ par molécule d'ATP). En conclusion, si l'on considère les 3 H+ nécessaires au fonctionnement de l'ATP synthase, ce sont 4 H+ qui entrent dans dans la matrice mitochondriale par molécule d'ATP synthétisée. |
| 8. Liens Internet |
|
"Plant Physiology Online" Liste de sites consacrés à l'ATP synthase. Notamment une animation fascinante sur la synthèse de l'ATP : "Animated movie of H+ pumping and ATP synthesis". |
![]()
{kind=link}