| Sumoylation et protéines SUMO ("Small Ubiquitin-like Modifier") |
| Tweet |
|
|
1. Introduction 2. Cibles des protéines SUMO 3. Mécanisme de la sumoylation |
4. Structure des protéines SUMO 5. Liens Internet et références bibliographiques |
|
1. Introduction
La fixation de protéines SUMO ("Small Ubiquitin-related MOdifier") à certaines protéines cibles est une modification post-traductionnelle avec des particularités :
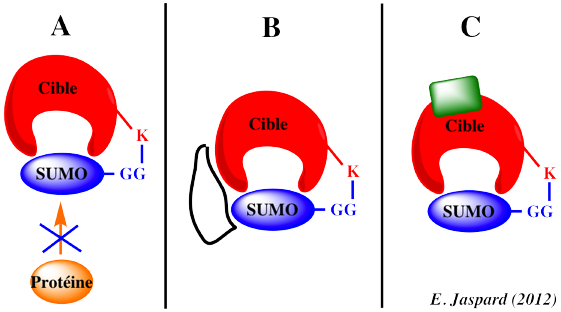
La sumoylation s'effectue via une cascade enzymatique analogue à l'ubiquitinylation. En revanche, le devenir des protéines sumoylées n'est pas la dégradation. La sumoylation peut induire 3 effets (non-mutuellement exclusifs) sur la protéine cible :
(A) Elle peut empêcher les interactions entre la protéine cible et une tierce protéine qui interagirait avec elle. Exemple : PCNA ("Proliferating Cell Nuclear Antigen") / Eco1. (B) Elle peut créer un nouveau site de fixation sur la protéine cible et induire de nouvelles interactions SUMO-dépendantes ("recruiting-factor"). Exemple : recombinaison au cours de la phase S - sumoylation de PCNA / hélicase Srs2. (C) Elle peut induire un changement de conformation de la protéine cible altérant ainsi son activité ou dévoilant un site de fixation jusque là masqué. |
|
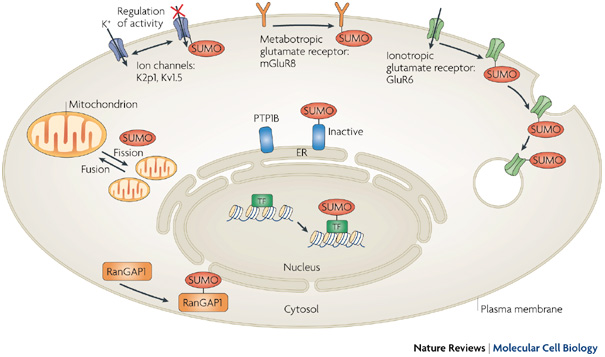
Un grand nombre de protéines cibles des protéines SUMO sont des protéines nucléaires (facteurs de transcription ou protéines impliquées dans l'organisation de la chromatine, la réplication et la réparation de l'ADN). Cependant, des cibles sont présentes dans le cytoplasme, la membrane plasmique, le réticulum endoplasmique et la mitochondrie. Figure ci-dessous : exemples de cibles des protéines SUMO.
Source : Geiss-Friedlander & Melchior (2007) La sumoylation :
|
Exemples de processus physiologiques dans lesquels la sumoylation joue un rôle très important (liste non exhaustive)
|
| Processus biologique | Protéine cible de SUMO | Fonction | Mécanisme | Organisme |
| Cytokines | Cdc3, 11, Shs1 | Désassemblage des anneaux septines | Pas élucidé | Levure |
| Canal ionique | KCNA5 | Modulation de l'activité | Pas élucidé | Mammifères |
| Activité des récepteurs | GLUK2 TbRI |
Internalisation Augmentation de l'activité |
Pas élucidé Recrutement de Smad3 |
Mammifères Mammifères |
| Transduction du signal | Smad4 | Modulation du signal | Pas élucidé | Mammifères |
| Transport | RanGAP1 | Translocation | Association à RanBP2 | Mammifères |
| Sous-compartimentation | PML | Formation des corps PML Dégradation des chaînes poly-sumoylées |
Interaction SUMO - "SUMO-Interacting-Motif" STUbL (RNF4) |
Mammifères |
| Biogénèse des ribosomes | Nop7 | Biogénèse des ribosomes et export | Pas élucidé | Levure |
| Réplication de l'ADN | PCNA | Inhibition de la recombinaison | Recrutement de Srs2 | Levure |
| Transcription | Mot1 Sp3 |
Degradation médiée par STUbL Répression de la transcription |
STUbL (SIx5/SIx8) Recrutement de méthyltransférases des histones |
Levure Mammifères |
| Réparation des cassures double-brin | BRCA1, 53BP1 Rad52 |
Réparation des cassures double-brin Régulation de l'activité Rad52 |
Augmentation de l'activité ligase Stabilisation, localisation |
Mammifères Levure, Mammifères |
| Réparation de l'excision des bases | TDG | Relarguage de l'ADN | Changement conformationnel | Mammifères |
| Mitose | Ndc10, Ndc80 Aurora B |
Mitose adéquate | Changement de localisation | Levure Mammifères |
| Méiose | Red1 | Formation de complexe synaptonemal | Fixation au complexe synaptonemal Zip1 | Levure |
| Maintien des télomères | TRF1, TRF2 | Homéostasie de la longueur du télomère | Recrutement de des corps nucléaires PML | Mammifères |
| Source : Creton & Jentsch (2010) Cell 143 | ||||
Un modèle mammifère, la souris, a été développé (Tirard et al., 2012) pour l'étude in vivo de la SUMOylation, qui est délicate techniquement.
|
|
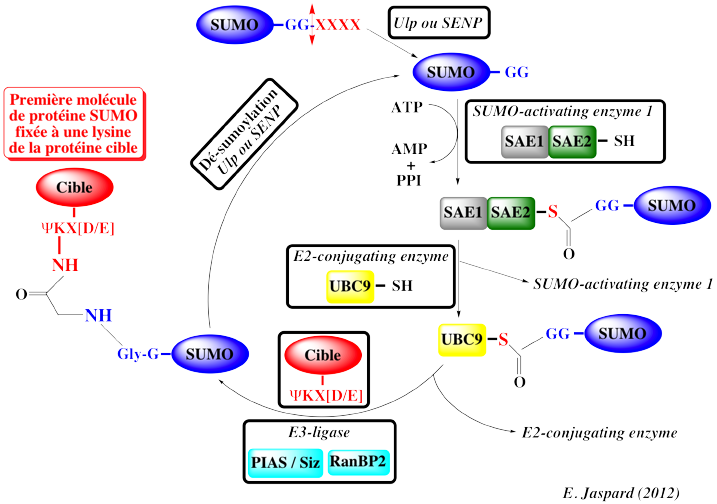
3. Mécanisme de la sumoylation La sumoylation résulte d'une cascade de réaction enzymatiques (comme l'ubiquitinylation). Les protéines SUMO sont synthétisées sous forme de précurseur inactif. 1ère étape : une protéase des protéines SUMO, "Ubiquitin-like Specific Protease" (Ulp) chez la levure ou "SENtrin-specific Protease" (SENP) chez l'homme clive 4 acides aminés de l'extrémité C-terminale du précurseur, ce qui fait apparaître une extrémité Gly-Gly. 2ème étape : la protéine SUMO est activée par "E1-activating enzyme" (avec hydrolyse de l'ATP), constituée de 2 sous-unités :
3ème étape : réaction de trans-estérification du résidu catalytique Cys de "E2-conjugating enzyme" - UBC9.
4ème étape : UCB9 forme une liaison isopeptidique entre la Gly C-terminale de la protéine SUMO et le groupement ε-aminé d'un résidu lysine (K) de la protéine cible. Le résidu lysine (en position 11 chez SUMO2 et SUMO 3), fait partie d'un motif consensus de sumoylation ("SUMO-Interacting-Motif" - SIM) : ψKX[D/E], où ψ est un gros résidu d'acide aminé hydrophobe (I, L ou V), X est n'importe quel acide aminé et [D/E] est un acide aminé acide. Chez certaines protéines cibles, un motif additionnel dans cette séquence consensus permet le couplage de la sumoylation à d'autres modifications post-traductionnelle. Le motif "Phosphorylation-Dependent SUMO Motif" - PDSM en est un exemple dans le cas de la phosphorylation : ψKX(D/E)XXSP avec une Ser qui peut être phosphorylée. La phosphorylation augmente le taux de "conjuguaison" de ces protéines cibles avec les protéines SUMO : le signal de phosphorylation est converti en un signal de sumoylation. 4ème étape (selon les protéines cibles) : la sumoylation de cibles spécifiques nécessite une "E3-ligase".
|
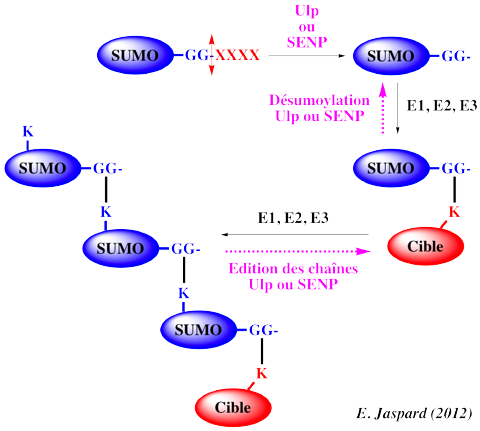
5ème étape : la désumoylation ("SUMO deconjugation") est essentielle à la régulation de ce processus. La même famille de protéine Ulp ou SENP catalyse :
Chez l'homme, 6 isoformes de SENP ont été identifiées et classées en 3 sous-familles : |
| Protéines de la voie de sumoylation | ||
| Protéine | Levure (Saccharomyces cerevisiae) | Mammifères |
| Modification ("modifier") | Smt3 | SUMO1 à SUMO4 |
| SUMO-activating enzyme subunit 1 (E1) | Aos1 / Uba2 | SAE1 / SAE2 |
| SUMO-conjugating enzyme (E2) / E.C. 6.3.2. | UBC9 | |
| SUMO-protein ligase (E3) / E.C. 6.3.2. |
|
|
| Désumoylation ("deconjugation") / E.C. 3.4.22.68 | Ulp1, Ulp2 | SENP1 à SENP7 |
|
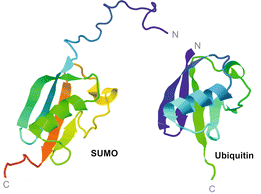
4. Structure des protéines SUMO Les protéines SUMO sont des protéines d'environ 100 - 115 acides aminés (10 - 12 kDa). Motif consensus de sumoylation - SIM ("SUMO-Interacting-Motif") L'analyse des séquences en acides aminés de ces motifs a permis d'établir, au minimum, 3 classes de SIM.
Les protéines SUMO sont des protéines similaires à l'ubiquitine. Les protéines SUMO et les ubiquitines n'ont environ que 20% d'homologie de séquences en acides aminés mais leur repliement sont trés semblables. Voir la protéine SUMO1 de l'homme : PDB 1A5R et l'ubiquitine de l'homme : PDB 1UBQ.
Source : Castro et al., 2012 |
|
Visualisation du complexe SUMO1 - UBC9 de Homo sapiens à une résolution de 2,4 Å Code PDB : 2PE6
L'interface SUMO-UBC9 non covalente semble importante pour :
|
| 5. Liens Internet et références bibliographiques |
|
Qiagen : "RNAi pathway" SUMOplot™ Analysis Program |
|
Geiss-Friedlander & Melchior (2007) "Concepts in sumoylation: a decade on" Nat. Rev. Mol. Cell Biol. 8, 947 - 956 Wilkinson & Henley (2010) "Mechanisms, regulation and consequences of protein SUMOylation" Biochem. J. 428, 133 - 145 |
|
Castro et al. (2012) "SUMO, a heavyweight player in plant abiotic stress responses" Cell Mol. Life Sci. 69, 3269 - 3283 Tirard et al. (2012) "In vivo localization and identification of SUMOylated proteins in the brain of His6-HA-SUMO1 knock-in mice" PNAS 109, 21122 - 21127 |
|
Capili & Lima (2007) "Structure and analysis of a complex between SUMO and Ubc9 illustrates features of a conserved E2-Ubl interaction" J. Mol. Biol. 369, 608 - 618 Hecker et al. (2006) "Specification of SUMO1- and SUMO2-interacting Motifs" J. Biol. Chem. 281, 16117 - 16127 |
![]()