| Mécanisme d'activation du récepteur de la thrombine (RCPG) |
| Tweet |
|
|
Le récepteur de la thrombine (PAR1 - "Protease-Activated Receptor") est une protéine transmembranaire de 425 acides aminés. La séquence partielle N-terminale extracellulaire du récepteur est la suivante : ATLDP40 RSFLL45 RNPND50 La thrombine est une enzyme qui hydrolyse entre
autre la protéine C et la protéine S aux sites suivants : En comparant la séquence clivée des substrats de la thrombine avec le fragment N-terminal de son récepteur, que peut-on suggérer ? |
|
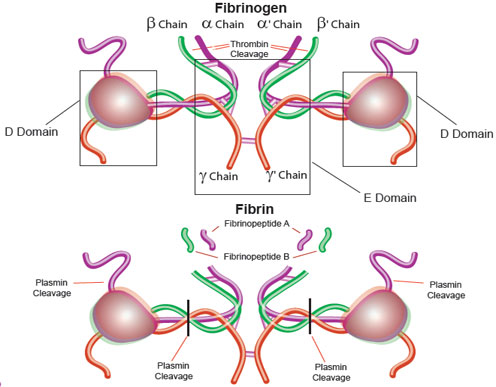
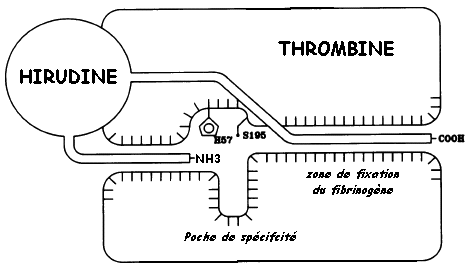
Dans la séquence N-terminale du récepteur, on remarque le site potentiel de clivage par la thrombine : ATLDPR41 / SFLL45 Recepteur KATNATLDPR SFLLRNPND ProtS ..TNAYPDLR SC....... ProtC ..QKDQLDPR IV....... Consensus ..tnaqlDpR sv....... Le fait que l'hirudine ne possède par d'arginine empêche qu'elle soit clivée par la thrombine qui hydrolyse la liaison peptidique après Arg et Lys. La thrombine protéolyse spécifiquement la liaison Arg / Gly du fibrinogène, libérant le fibrinopeptide A. La perte de ce petit peptide n'est cependant pas suffisante pour rendre insoluble la molécule de fibrine résultante.
Source : Sigma La thrombine hydrolyse alors un second peptide de la fibrine : le fibrinopeptide B. Les monomères de fibrine polymérisent alors spontanément pour former un gel insoluble. L'activation du récepteur de la thrombine est testée avec les composés suivants :
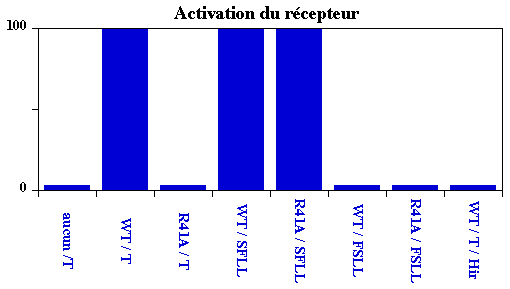
Les résultats obtenus sont présentés dans la figure ci-dessous :
Sachant que le fragment Hir de l'hirudine et que la séquence YEPF WEDEE60 du récepteur se fixent de manière réversible à la thrombine, que peut-on conclure sur le mécanisme probable d'activation du récepteur ? |
| Récepteur | Thrombine ou peptide | Activation | Interprétation |
| aucun | thrombine | 0 | Témoin négatif. |
| WT : LDPR41 / SFLL45 | thrombine | 100 | Témoin positif. |
| R41A : LDPA41 / SFLL45 | thrombine | 0 |
Pas d'hydrolyse N-terminale du récepteur ? |
| WT | SFLL | 100 |
Le peptide de synthèse SFLL
mime celui qui apparait quand il y a coupure du récepteur ==>
S est indispensable pour l'autoactivation
du récepteur. |
| R41A | SFLL | 100 | Donne la réponse aux questions ci-dessus : R41 est indispensable pour que l'extrémité N-terminale du récepteur soit coupée par la thrombine. |
| WT | FSLL | 0 | Confirme que S est indispensable à cette position pour l'autoactivation du récepteur. |
| R41A | FSLL | 0 | Confirme que S est indispensable à cette position pour l'autoactivation du récepteur. |
| WT | thrombine + Hir | 0 |
Confirme que c'est l'extrémité C-terminale de l'hirudine (DFEEI PEEYL) qui est indispensable pour l'inactivation (irréversible) de la thrombine.
|
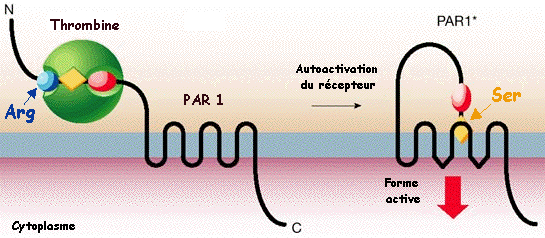
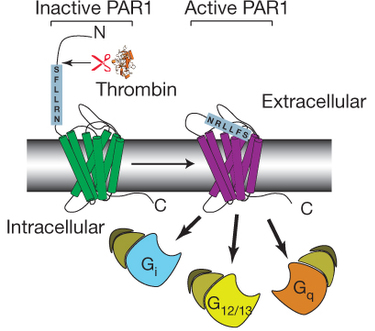
Mécanisme de l'autoactivation de PAR1 par la thrombine La thrombine (sphère verte) hydrolyse un peptide (R41 / SF) près de l'extrémité N-terminale de son récepteur PAR1. La nouvelle extrémité N-terminale du récepteur SF interagit avec son domaine transmembranaire. Cette interaction modifie la conformation du récepteur ce qui l'active.
Source : Coughlin (2000) Le récepteur de la thrombine PAR1 ("Protease-Activated Receptor") est un récepteur couplé à une protéine G (RCPG). Dans les plaquettes humaines, PAR1 (couplé à une protéine G de la classe Gi/o) active la phosphoinositide 3-kinase (PI3K). Les phosphoinositides sont des dérivés phosphorylés du phosphatidylinositol. L'activation de PI3K régule l'activation de l'intégrine αIIbβ3 plaquettaire et donc l'agrégation plaquettaire et potentialise l'augmentation de la concentration de calcium intra-plaquettaire.
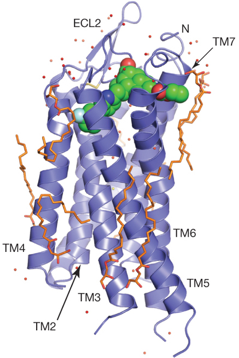
Source : Zhang et al. (2012) Figure ci-dessous : structure de PAR1. Précurseur de PAR1 : 425 acides aminés; acides aminés 1 à 26 : peptide signal; acides aminés 27 - 41 : propetide clivé pour l'activation du récepteur.
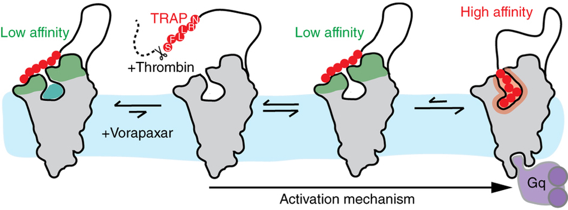
Source : Zhang et al. (2012) Le vorapaxar (SCH 530348 - molécule en vert) est un antagoniste de PAR1, basé sur un alcaloïde naturel, l'himbacine. Modèle de fixation du peptide SFLLRN sur PAR1 L'étude de la courbe [force - distance] basée sur la microscopie de force atomique permet de quantifier la force de liaison dynamique de RCPG dans la membrane avec le peptide SFLLRN ("Thrombin Receptor-Activating Peptide" - TRAP) et le vorapaxar. Le peptide (TRAP - figure ci-dessous) se fixe avec une faible affinité, probablement via les boucles 2 et 3 extracellulaires, aussi bien dans le cas de PAR1 que de PAR1 inhibée par le vorapaxar (PDB : 3VW7).
Source : Alsteens et al. (2015)
|
| Liens Internet et références bibliographiques |
|
Vu et al. (1991) "Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation" Cell 64, 1057 - 1068 Coughlin (2000) "Thrombin signalling and protease-activated receptors" Nature 407, 258 - 264 |
|
|
De Candia ("2012) "Mechanisms of platelet activation by thrombin: A short history" Thromb. Res. 129, 250 - 256 Zhang et al. (2012) "High-resolution crystal structure of human protease-activated receptor 1" Nature 492, 387 - 392 Alsteens et al. (2015) "Imaging G protein–coupled receptors while quantifying their ligand-binding free-energy landscape" Nat. Met. 12, 845 - 851 |
Synthèse de cette correction au format PDF (tous droits réservés biochimej). |
![]()