| Les pseudoenzymes : homologues catalytiquement inactifs d'enzymes |
| Tweet |
|
|
1. Introduction 2. Découverte des pseudoenzymes 3. Exemples de pseudoenzymes et de familles d'enzymes 4. Différents modes de fonctionnement des pseudoenzymes |
5. Conservation des résidus catalytiques et contrainte évolutive à long terme 6. Vitesse d'évolution des sites critiques pour la relation structure - fonction 7. L'annotation et la découverte de pseudoenzymes 8. Liens Internet et références bibliographiques |
1. Introduction Au cours de l'évolution des enzymes, la duplication des gènes et l'accumulation de mutations se sont traduites par la perte de résidus d'acides aminés impliqués dans la catalyse et/ou la fixation du substrat ou par un changement de la spécificité du substrat ou de la réaction catalysée. Cette évolution a donné naissance à un groupe de protéines, liées aux enzymes, qui ont perdu leur pouvoir catalytique : les pseudoenzymes. Cependant, la structure tridimensionnelle de l'enzyme d'origine est globalement conservée chez la pseudoenzyme : cette dernière possède donc une nouvelle fonction (non catalytique), souvent liée à la régulation de l'activité de l'enzyme originelle.
Les pseudoenzymes sont universelles :
Les pseudoenzymes sont différentes des protéines moonlighting. |
2. Découverte des pseudoenzymes Le premier exemple d'un homologue non catalytique d'une enzyme date de 1967 quand on a découvert que la séquence partielle de l'α-lactalbumine bovine a une identité de séquence d'environ 35% avec le lysozyme de poulet :
Cette similarité pouvait être causée par une évolution convergente ou divergente : un gène ancestral semblable à un lysozyme avait été dupliqué et l'un de ses descendants avait été muté en un gène codant pour la forme non enzymatique, α-lactalbumine. Les pseudoenzymes ont des fonctions essentielles dans la signalisation et le métabolisme. Parmi les exemples importants d'homologues inactifs, on peut citer entre autres, les pseudoenzymes liées aux protéases, aux kinases, aux phosphatases, aux enzymes de conjugaison de l'ubiquitine E2 et aux phospholipases. |
| Exemples de pseudoenzymes | ||
| Nom | Famille de l'enzyme | Rôle et relation avec l'homologue actif (si celui-ci est défini) |
| CPSF100 (homme), Cft2 (levure) | Endonucléase | Sous-unité du complexe CPSF impliquée dans le clivage des précurseurs d'ARN messagers - le complexe comprend sa nucléase active apparentée CPSF73 (homme) ou Ysh1 (levure) |
| cFLIP (vertébrés) | Caspase (protéase à cysteine) | Hétérodimérise avec la caspase 8 et inhibe l'activation de la caspase - régule l'activation de NF-κB |
| Tre1, Tre2 (levure) | Liée au récepteur de la transferrine (elle-même une métallo-carboxypeptidase inactive) | Tri et dégradation des récepteurs transmembranaires endocytés |
| KSR (metazoaires) | Kinase | Echafaudage pour d'autres kinases actives - contrôle de la propagation du signal |
| BMSPH1 (Bombyx mori) | Protéase à sérine | Mélanisation des nodules à la suite d'une infection bactérienne - se lie à une lectine qui reconnaît les bactéries Gram- |
| Protéases inactives paralogues chez les acariens de la gale (multiple) | Protéase à sérine, protéase à cysteine | Evasion immunitaire, éventuellement par inhibition du système du complément |
| Scarface (mouche) | Protéase à sérine | Régulateur négatif sécrété de la voie JNK |
| EPHB6 (vertébrés) | EPH (RTK) | Développement du système immunitaire, prolifération des cellules T |
| Composants ARN exosome (multiples) | Ribonucléase | Complexe contenant des nucléases actives et leurs composés apparentés inactifs - Traitement et dégradation des ARN |
| Homologues ornithine décarboxylase (eucaryotes) | Ornithine décarboxylase | Biosynthèse des polyamines - régulation de l'activité de l'ornithine décarboxylase |
| ThnY (bactéries) | Ferredoxine réductase | Stabilise la liaison du facteur de transcription ThnR aux promoteurs de gènes |
| Famille MAGUK (multiple) | Guanylate kinase | Forme un échafaudages pour les complexes de signalisation - régulation de la signalisation |
| NDRG (vertébrés) | α-hydrolase, β-hydrolase | Suppresseur de tumeur, régulateur négatif de la voie de la β-caténine |
| XPC (homme), Rad4 (levure) | Transglutaminase | Réparation de l'ADN, excision des nucléotides - interaction protéine-protéine |
BMSPH1 : homologue 1 de la protéase de Bombyx mori - cFLIP : protéine inhibitrice de type FLICE - Cft2 : facteur de clivage 2 (également appelé Ydh1) - CPSF : facteur de spécificité de clivage et de polyadénylation - EPHB6 : récepteur 6 de l'éphrine de type B - JNK : kinase N-terminale Jun - KSR : suppresseur de kinase de RAS - NDRG : gène NMYC régulé en aval - NF-κB : facteur nucléaire κB - Tre : protéine semblable au récepteur de la transferrine - RTK : récepteur tyrosine kinase - XPC : protéine complémentaire du groupe C (maladie xeroderma pigmentosum). Source : Adrain & Freeman (2012) |
||
3. Exemples de pseudoenzymes et de familles d'enzymes La prévalence d'homologues enzymatiques catalytiquement inactifs est plutôt une règle qu'une exception : la plupart des familles d'enzymes possèdent des pseudoenzymes qui constitueraient environ 10% du protéome. Les pseudoenzymes sont souvent bien conservées, ce qui implique une pression sélective permanente pour les conserver et maintenir leur fonction. |
| Exemples d'homologues enzymatiques catalytiquement inactifs | ||
| Famille de pseudoenzyme | Uniprot | Rôle |
| Pseudophosphatase | O01767 | Pseudophosphatase de l'oeuf 4 de C. elegans. |
| Q86WG5 | Pseudophosphatases SBF2/MTMR13 liées à la maladie neurodégénérative de Charcot - Marie - Tooth. | |
Pseudokinase |
Environ 10% des protéines kinases humaines sont supposées inactives et classées comme pseudokinases car elles ne contiennent pas les résidus d'acides aminés du domaine kinase nécessaires à la catalyse. | |
| Q96MK3 | Pseudokinase FAM20A de l'homme. Elle agit comme activateur allostérique de la Ser/Thr protéine kinase FAM20C (appareil de Golgi) : le complexe formé augmente la capacité de FAM20C à phosphoryler les protéines qui forment la matrice qui guide le dépôt des minéraux de l'émail (biominéralisation des dents). | |
| Q87VB1 | La pseudokinase sélénoprotéine-O (SelO, hautement conservée - code PDB : 6EAC) transfère l'AMP des résidus ATP aux résidus Ser, Thr et Tyr sur des substrats protéiques (AMPylation) : il s'agit d'une nouvelle activité enzymatique de la superfamille des protéines kinases. | |
| Pseudoenzyme de désubiquitinylation | Q15018 | BRCC36 est une enzyme de désubiquitinylation des chaînes d'ubiquitine liées à K63. Son domaine MPN+ s'associe à la pseudoenzyme de désubiquitinylation KIAA0157 (appelée aussi Abraxas) : les contacts établis lors de l'association [BRCC36 / KIAA0157] sous forme de dimère d'hétérodimères permettent à BRCC36 de passer dans la conformation active et d'interagir avec ses protéines cibles. Ce mécanisme de "super-dimérisation" permet ainsi à cette pseudoenzyme de désubiquitinylation d'activer de manière allostérique son enzyme apparentée et de réguler sa localisation subcellulaire. |
| Pseudosynthase | Q9ZNR6 | Les plantes (exemple, Arabidopsis thaliana) stimulent l'expression de la pseudoenzyme PDX1.2 lors d'un stress thermique et au cours du développement embryonnaire. PDX1.2 régule positivement la production de vitamine B6 par hétéro-association avec ses homologues catalytiques actifs PDX1.1 et PDX1.3 ("Pyridoxine biosynthesis 1 & 2"). La structure de PDX1.2 adopte un repliement quasi-identique à celui de PDX1.3 et maintient toutes les interactions protéine-protéine nécessaires entre les sous-unités (code PDB : 6HX3). Cependant, il y a une inversion de la distribution de charges à la surface de PDX1.2 qui aurait un impact sur l'efficacité de rotation du site P2 voisin de PDX1.3. La pseudoenzyme PDX1.2 agirait comme un "module de réglage électrostatique" combiné à un mécanisme d'hétéro-association basé sur une incorporation aléatoire. |
| PseudoGTPase | Les protéines p190RhoGAP-A (ARHGAP35) et p190RhoGAP-B (ARHGAP5) sont des régulateurs clés de la signalisation Rho GTPase et sont essentielles à la structure et à la contractilité de l'actine du cytosquelette.
Source : Stiegler & Boggon (2017) p190RhoGAP-A et p190RhoGAP-B (code PDB : 5U4V) partagent plus de 50% d'identité de séquences et ont une organisation identique :
|
|
| Pseudoisomérase | O75344 | La peptidyl-prolyl cis-trans pseudoisomérase FKBP6 est une protéine co-chaperon (en interaction avec HSP90) nécessaire au cours de la spermatogenèse pour réprimer les éléments transposables (via des piARN et des protéines Piwi pendant la méiose) et empêcher leur mobilisation qui est essentielle à l'intégrité de la lignée germinale. Bien qu'elle contienne un domaine de type PPIase FKBP, elle ne possède pas d'activité peptidyl-prolyl cis-trans isomérase. |
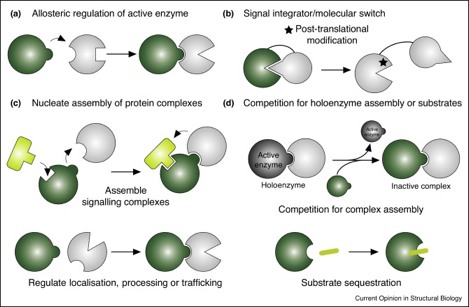
4. Différents modes de fonctionnement des pseudoenzymes Les pseudoenzymes (en vert, figure ci-dessous) exercent leurs effets sur la transduction du signal ou sur le métabolisme via des interactions avec d'autres protéines (notamment les enzymes "clientes", en gris) ou avec des substrats (en jaune).
Source : Murphy et al. (2017) (a) Régulation allostérique (positive ou négative) de l'activité catalytique d'une enzyme "cliente" (fréquemment, une enzyme apparentée). (b) Les domaines d'une pseudoenzyme peuvent subir une modification post-traductionnelle (indiquée par une étoile) comme l'ubiquitinylation, la (dé)-phosphorylation ou le clivage protéolytique, susceptible de faire basculer la structure de la pseudoenzyme dans une conformation ayant une autre fonction. (c) En jouant un rôle de "domaine d'interactions protéiques", les pseudoenzymes peuvent servir de "noyau" d'assemblage des sous-unités constitutives de complexes protéiques pour rapprocher une enzyme de son substrat (en haut) ou réguler la localisation des protéines, leur stabilité ou le contrôle qualité (en bas). (d) Les pseudoenzymes peuvent entrer en compétition avec leurs enzymes canoniques actives et, ainsi, empêcher l'assemblage de complexes protéiques (en haut) ou séquestrer les molécules de substrat, dès lors indisponibles pour la réaction enzymatique (en bas). |
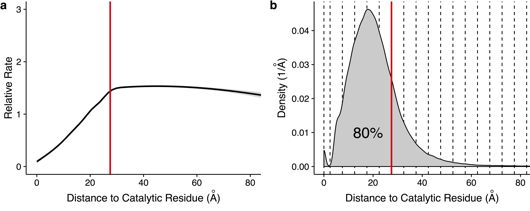
5. Conservation des résidus catalytiques et contrainte évolutive à long terme Les résidus d'acides aminés des sites catalytiques sont remarquablement conservés. Le site catalytique des enzymes induit une contrainte évolutive à long terme :
Source : Jack et al. (2016) Figure a, ci-dessus : Taux d'évolution spécifique d'un site en fonction de la distance avec le résidu catalytique le plus proche. La courbe représente le taux moyen pour tous les résidus d'acides aminés, calculé à partir d'un modèle additif généralisé. Les taux augmentent presque linéairement avec la distance jusqu'à 27,5 Å (ligne verticale). Figure b, ci-dessus : Diagramme de densité des résidus avec une distance donnée par rapport à un résidu catalytique. La ligne rouge verticale indique une distance de 27,5 Å et 80% des résidus se situent en dessous de cette distance. La comparaison des propriétés évolutives des enzymes avec celles de leur homologue pseudoenzyme (qui adoptent le même repliement tridimensionnel) indique que la contrainte évolutive à long terme chez les enzymes est significativement réduite chez les pseudoenzymes, malgré une similarité structurale très élevée (RMSD ≈ 1,5 Å en moyenne). Cette réduction significative est observée également avec les pseudoenzymes qui conservent la capacité de fixer les ligands de leur homologue enzyme.
|
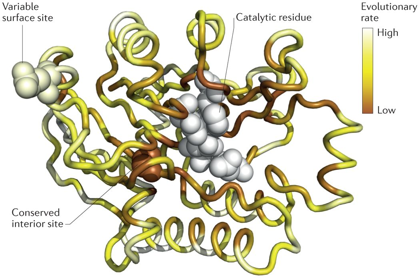
6. Vitesse d'évolution des sites critiques pour la relation structure - fonction La vitesse d'évolution peut être défini comme le nombre de substitutions (mutations fixes) par unité de temps au cours de l'évolution des séquences des protéines. Les contraintes structurales et fonctionnelles régissent la vitesse d'évolution des sites critiques pour la relation structure - fonction des protéines :
Source : Echave et al. (2016) Les modèles biophysiques (basés en particulier sur l'accessibilité au solvant et la compacité locale) indiquent que la vitesse d'évolution spécifique d'un site est liée au changement de stabilité thermodynamique qu'induit une mutation donnée. Cependant, les modèles actuels n'expliquent (au plus) qu'environ 60% de la variation observée de la vitesse d'évolution spécifique d'un site. |
7. L'annotation et la découverte de pseudoenzymes a. L'annotation des pseudoenzymes La base de données UniProtKB est un recueil d'informations sur les protéines : plus de 150 millions de protéines y sont recensées. Dans le cas des enzymes (qui représentent 20 à 40% de la plupart des protéomes) :
Cependant, si l'annotation des enzymes est bien définie, celle des pseudoenzymes est plus délicate :
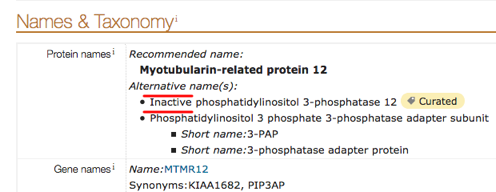
Dans le champ de recherche de l'interface web de UniProt (en haut de la page), le terme de recherche "inactive" dans le champ "Protein name" permet de récupérer des fichiers de pseudoenzymes.
En ce qui concerne les mots-clé de l'ontologie :
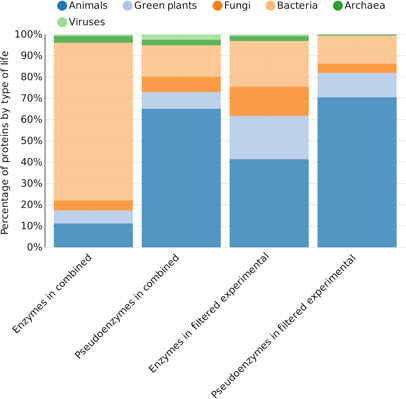
Voir l'ensemble des fichiers des homologues Swiss-Prot/EMBL-EBI avec ou sans preuves expérimentales de la fonction, où Les pseudoenzymes ont été identifiées sur la base de l'absence d'annotation de la catalyse ou de l'absence explicite d'annotations de fonction ("Combined, Experimental, Filtered Combined, Filtered Experimental"). La figure ci-dessous montre la répartition des enzymes et des pseudoenzymes selon les espèces pour les ensembles de données "Combined" et "Filtered Experimental" :
Source : Ribeiro et al. (2019b) |
b. La découverte de pseudoenzymes Les critères actuels pour identifier les pseudoenzymes découlent des preuves de l'activité catalytique chez leurs homologues enzymes actives. En d'autres termes, la prédiction qu'une protéine est une pseudoenzyme dépend de la connaissance du mécanisme catalytique et des résidus impliqués des membres catalytiquement actifs de la famille à laquelle cette pseudoenzyme appartient. Il existe des pseudoenzymes qui ont conservé le site actif et, par ailleurs, la mutation d'un résidu catalytique n'abolit pas forcément l'activité catalytique.
L'alignement des séquences d'acides aminés des protéines est principalement utilisée pour prédire l'absence de résidus d'acides aminés dont on a démontré expérimentalement qu'ils sont indispensables à l'activité enzymatique.
Source : Gnangnon et al. (2019)
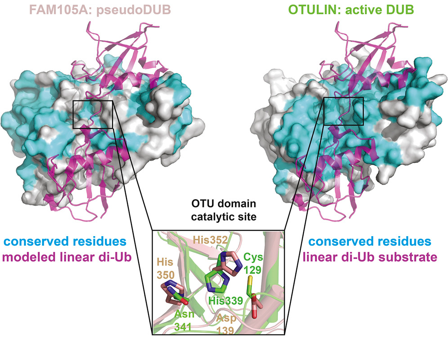
La comparaison des données structurales (quand on en dispose) d'une enzyme et de sa pseudoenzyme permet de décrire :
Source : Ceccarelli et al. (2019) La pseudoenzyme [FAM105A / OTULINL] est une pseudodésubiquitinase de la famille "Ovarian Tumor Protease" (OTU). Le domaine OTU présente des déficiences structurales du site actif et du site de fixation du substrat (Cys 139 du site actif remplacée par Asp 139) responsables de l'absence d'activité catalytique vis-à-vis des liaisons avec l'ubiquitine et de l'incapacité de FAM105A à se lier à l'ubiquitine contrairement à l'enzyme [FAM105B / OTULIN] catalytiquement active. La mise au point d'outils de prédiction fiables permettant d'identifier et d'annoter correctement les pseudoenzymes nécessite d'accroître les connaissances des mécanismes de toutes les réactions enzymatiques. A cela s'ajoute le développement de recueils de données tels que :
Enfin, sur le plan théorique, l'augmentation des performances des logiciels de modélisation moléculaire des protéines et des méthodes d'apprentissage profond (intelligence artificielle) seront déterminantes. |
| 8. Liens Internet et références bibliographiques |
ProteinPseudokinaseDomain : collection annotée des séquences de pseudokinase prédites et de leurs relations évolutives Identification de pseudoenzymes potentiels dans CATH via les domaines structuraux The CATH Protein Structure Classification EMBO : Pseudoenzymes 2018 conference |
|
Brew et al. (1967) "Comparison of the amino acid sequence of bovine alpha-lactalbumin and hens egg white lysozyme" J. Biol. Chem. 242, 3747 - 3749 |
Article |
Adrain & Freeman (2012) "New lives for old: evolution of pseudoenzyme function illustrated by iRhoms" Nat. Rev. Mol. Cell. Biol. 13, 489 - 498 Zeqiraj et al. (2015) "Higher -Order Assembly of BRCC36–KIAA0157 Is Required for DUB Activity and Biological Function" Mol. Cell. 59, 970 - 983 |
|
Jack et al. (2016) "Functional Sites Induce Long-Range Evolutionary Constraints in Enzymes" PLoS Biol. 14, e1002452 Echave et al. (2016) "Causes of evolutionary rate variation among protein sites" Nat. Rev. Genet. 17, 109 - 121 |
|
Stiegler & Boggon (2017) "p190RhoGAP proteins contain pseudoGTPase domains" Nat. Commun. 8, 506 Murphy et al. (2017) "Live and let die: insights into pseudoenzyme mechanisms from structure" Curr. Opin. Struct. Biol. 47, 95 - 104 Eyers et al. (2017) "Tribbles in the 21st Century: The Evolving Roles of Tribbles Pseudokinases in Biology and Disease" Trends Cell Biol. 27, 284 - 298 Lisa et al. (2017) "The crystal structure of PknI from Mycobacterium tuberculosis shows an inactive, pseudokinase-like conformation" FEBS J. 284, 602 - 614 |
|
Sreelatha et al. (2018) "Protein AMPylation by an Evolutionarily Conserved Pseudokinase" Cell 175, 809 - 821 Sharir-Ivry & Xia (2018) "Nature of Long-Range Evolutionary Constraint in Enzymes: Insights from Comparison to Pseudoenzymes with Similar Structures" Mol. Biol. Evol. 35, 2597 - 2606 |
|
Ribeiro et al. (2019a) "Emerging concepts in pseudoenzyme classification, evolution, and signaling" Sci. Signal. 594, eaat9797 Kwon et al. (2019) "Tracing the origin and evolution of pseudokinases across the tree of life" Sci. Signal. 12, pii: eaav3810 Novikova et al. (2019) "The tunable intrahexamer heteroassembly mechanism of pseudoenzyme/enzyme pair, PDX1.2/PDX1.3, with its molecular impact on vitamin B6 regulation" bioRxiv doi.org/10.1101/717082 |
|
Zaru et al. (2019) "Challenges in the annotation of pseudoenzymes in databases: the UniProtKB approach" FEBS J. doi: 10.1111/febs.15100 Gnangnon et al. (2019) "Plasmodium pseudo-Tyrosine Kinase-like binds PP1 and SERA5 and is exported to host erythrocytes" Sci. Rep. 9, 8120 Robinson et al. (2019) "Crystal structure of the pseudoenzyme PDX1.2 in complex with its cognate enzyme PDX1.3: a total eclipse" Acta Crystallogr. D Struct. Biol. 75, 400 - 415 |
|
Ribeiro et al. (2019b) "Identifying pseudoenzymes using functional annotation. How loss of function correlates with mutations in the catalytic site" FEBS J. doi: 10.1111/febs.15142 Ceccarelli et al. (2019) "FAM105A/OTULINL Is a Pseudodeubiquitinase of the OTU-Class that Localizes to the ER Membrane" Structure 27, 1000 - 1012 |
![]()