| Les protéines à plusieurs fonctions ou protéines "moonlighting" |
| Tweet |
|
|
1. Introduction 2. Exemples de protéines moonlighting 3. Illustrations de différents rôles de quelques protéines moonlighting a. Enzymes du métabolisme |
d. Les lysines méthyltransférases et les lysines déméthylases 4. Protéines moonlighting et liaison à l'ARN 5. Activités dans le métabolisme et fonctions dans le noyau 6. Identification de protéines moonlighting 7. Liens Internet et références bibliographiques |
1. Introduction Les protéines constituent la classe de bio-macromolécules la plus polyvalente. L'un des aspects de cette polyvalence réside dans la capacité qu'ont certaines protéines d'avoir des fonctions différentes, sans lien. De multiples fonctions pour des protéines augmente la complexité du fonctionnement de la cellule et ne peut que lui être bénéfique. De telles protéines ont été mises en évidence pour la première fois par Piatigorsky et collaborateurs : ils ont montré que les protéines δ-crystallin et argininosuccinate lyase (EC 4.3.2.1) de canard sont codées par le même gène, phénomène qu'ils ont appelé partage de gènes ("gene sharing") :
Dénomination protéines "moonlighting" Le terme "partage de gènes" est cependant ambigü puisqu'un gène peut coder pour différentes protéines (différents sites d'initiation de la traduction ou de la terminaison, épissage alternatif, ...). La dénomination protéines moonlighting ("moonlighting proteins", traduit par "protéines au noir") a été proposée par C. Jeffery en 1999, par analogie avec "travail au noir" (un second emploi en plus d’un emploi principal). Plus récemment, des termes plus génériques ont été employés pour décrire ces protéines : protéines multi-tâches ("multitask proteins") et protéines multi-fonctionnelles extrêmes ("extreme multifunctional proteins"). Les protéines moonlighting sont différentes des pseudoenzymes. Importance de l'étude des protéines moonlighting
|
2. Exemples de protéines moonlighting Le groupe des protéines moonlighting incluent les protéines sécrétées en raison de de leurs nombreux rôles dans différents organismes. En revanche, il n'inclue pas :
|
| Exemples de protéines moonlighting | |||
| Protéine | Fonction canonique ou voie métabolique | Fonction moonlighting | Organisme |
| Hexokinase | Glycolyse | Régulation de la transcription | S. cerevisiae |
| Signalisation du glucose - contrôle de la mort cellulaire | Arabidopsis thaliana | ||
| Phosphofructokinase | Glycolyse | Autophagie | P. pastoris |
| Aldolase | Glycolyse | Assemblage de l'ATPase H+ vacuolaire | S. cerevisiae |
| Enolase | Glycolyse | Fusion vacuolaire - import d'ARNt mitochondrial | |
| Homocitrate synthase | Acyl transférase (biosynthèse de la lysine ) | Réparation de l'ADN endommagé | |
| Superoxide dismutase | Conversion du radical superoxyde en O2 et H2O2 | Régulation des gènes liés au stress oxydatif | |
| Pyruvate carboxylase | Reconstitution du cycle du TCA | Assemblage de l'alcool oxydase | H. polymorpha |
| Aconitase | Cycle de Krebs | Homéostasie du fer | Homo sapiens |
| Clathrine | Traffic membranaire | Stabilité du fuseau mitotique | |
| Galactokinase | Métabolisme du galactose | Régulation de la transcription | S. cerevisiae, K. lactis |
| Zuotine | Chaperone | Activation de la résistance aux médicaments pléiotropes | S. cerevisiae |
| Arginase | Hydrolyse de l'arginine | Inhibiteur de l'ornithine transcarbamylase | |
| Inositol phosphate kinase | Biosynthèse de l'inositol | Régulation de la transcription | |
| Acétohydroxyacide réducto-isomérase | Synthèse d'acides aminés à chaîne ramifiée | Entretien de l'ADN mitochondrial | |
| Sous-unité SDH3 de la succinate déshydrogénase | Oxydation du succinate (cycle de l'acide citrique) - transport d'électrons | Assemblage de la translocase mitochondriale | |
| Enolase | Déshydratation du 2-phosphoglycérate (glycolyse) | Liaison au plasminogène et/ou aux régulateurs du complément | C. albicans |
| Phosphoglycérate mutase | Isomérisation du 3-phosphoglycérate (glycolyse) | ||
| Glycérol 3-phosphate déshydrogénase | Réduction de la dihydroxyacétone phosphate en glycérol 3-phosphate (auxiliaire de la glycolyse) | ||
| HGT1 | Transporteur du glucose | ||
| HAL3 | Décarboxylation de la cystéine phosphate pantothényl (biosynthèse du CoA) | Inhibition des phosphatases PPZ 1 et 2 | S. cerevisiae, S. pombe |
| Thiorédoxine | Antioxydant | Sous-unité de l'ADN polymérase du bactériophage T7 | E. coli |
Sources : Flores & Gancedo (2011), Gancedo et al. (2016) Voir une liste des numéros d'accession Uniprot d'un grand nombre de protéines moonlight. |
|||
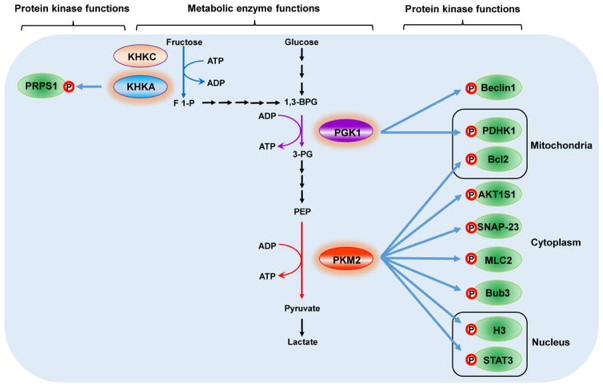
3. Illustrations de différents rôles de quelques protéines moonlighting Certaines enzymes du métabolisme telles que l'hexokinase, la phosphoglycérate kinase 1, la pyruvate kinase M2, l'isoforme A de la céto-hexokinase et les nucléosides diphosphates kinases 1 et 2 qui phosphorylent des métabolites solubles ont également une activité protéine kinase.
Source : Lu & Hunter (2018) Cette activité régule une grande diversité de processus tels que l'effet Warburg, la transcription des gènes, le cycle cellulaire, l'apoptose, l'autophagie, la sécrétion de l'exosome, l'activation des cellules T, le transport du fer, l'ouverture de canaux ioniques et d'autres fonctions cellulaires fondamentales. Voir également : Snaebjornsson & Schulze (2018). |
|
Le facteur d'élongation thermique instable Tu (EF-Tu) est une protéine G qui catalyse la fixation de l'aminoacyl-ARNt au site A du ribosome. Cependant, EF-Tu a d'autres fonctions à la surface des cellules eucaryotes et procaryotes. En effet, EF-Tu peut se déplacer vers la surface extracellulaire et y rester : il y interagit avec des récepteurs membranaires et avec la matrice extracellulaire. Des motifs structuraux linéaires courts ("short linear motifs", SLiMs) dans des régions non conservées exposées à la surface joueraient un rôle clé dans les fonctions moonlighting de cette protéine très abondante et ancienne. En particulier, un groupe de résidus d'acides aminés basiques situé à l'extrémité N-terminale est associé à des fonctions moonlighting de EF-Tu. Ce SLiM, situé dans une région désordonnée, contient au moins 3 résidus exposés à la surface et il est prédit pour établir des interactions protéine:protéine.
Source : Harvey et al. (2019) Les résidus Arg7 et Lys9 ne sont pas conservés chez certaines espèces bactériennes et pourraient résulter de mutations ponctuelles avantageuses. |
|
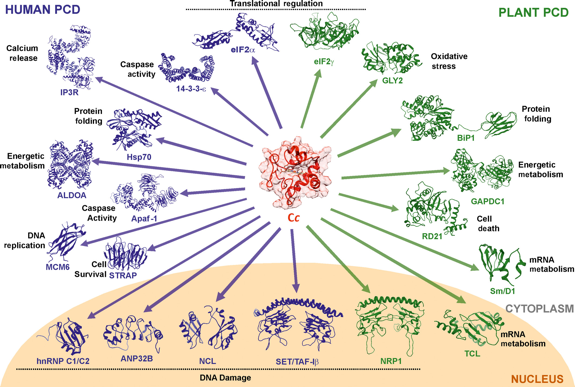
Le cytochrome c est un transporteur d'électrons de la chaîne respiratoire dans les mitochondries. Cependant, il joue aussi un rôle en dehors des mitochondries qui induit la transition des cellules apoptotiques de la vie vers la mort. En effet, lorsque des cellules sont endommagées, le cytochrome c est libéré dans le cytosol :
De nombreuses nouvelles cibles protéiques du cytochrome c ont été décrites. Notamment les chaperonnes des histones après des cassures de l'ADN : le cytochrome c pourrait ainsi participer à la modulation de la dynamique de la chromatine par la fixation compétitive à ces protéines chaperons. Figure ci-dessous : interactions avec le cytochrome c de protéines du cytoplasme ou du noyau de cellules humaines (en bleu) ou d'Arabidopsis thaliana (en vert).
Source : Gonzalez-Arzola et al. (2019) |
d. Les lysines méthyltransférases et les lysines déméthylases Presque toutes les enzymes qui ajoutent ou éliminent les groupes méthyle des résidus lysine ont été découvertes initialement en tant qu'enzymes modifiant les histones : elles ont donc d'abord été nommées histones lysines méthyltransférases et histones lysines déméthylases, respectivement. Au fur et à mesure que des preuves que ces enzymes agissent sur des protéines autres que les histones ont été apportées, elles ont été renommées lysines méthyltransférases et lysines déméthylases, afin de refléter leur rôle plus général dans la modification des lysines des protéines. Exemples : SETD7, G9a et SMYD2. La méthylation des lysines des histones et des protéines autres que les histones :
|
|
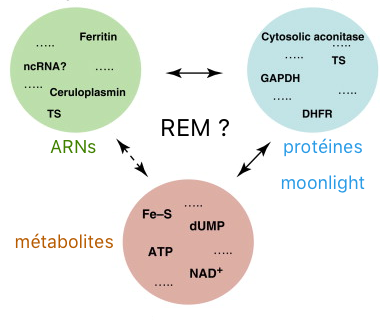
4. Protéines moonlighting et liaison à l'ARN Beaucoup d'enzymes moonlighting agissent comme protéines de liaison à l'ARN. Cependant, aucun concept global n'a encore été proposé pour ce phénomène. L'hypothèse de l'interaction [ARN - enzyme moonlighting - métabolite] ("RNA-enzyme-metabolite hypothesis", REM) suggère l'existence de "liens de régulation" entre la [transcription des gènes / traduction des ARN messagers] et le métabolisme intermédiaire : ces liens seraient contrôlés par les enzymes moonlighting du métabolisme qui se fixent à l'ARN.
Source : Hentze & Preiss (2010) Exemple de la glycéraldéhyde 3-phosphate déshydrogénase (GAPDH), une enzyme de la glycolyse dans le cytosol :
Exemple de l'aconitase de l'homme (ACO1), une enzyme du cycle de l'acide tricarboxylique :
|
5. Activités dans le métabolisme et fonctions dans le noyau De nombreuses enzymes du métabolisme localisées dans le cytoplasme (notamment toutes les enzymes essentielles de la glycolyse) et des enzymes mitochondriales ont également des fonctions de type moonlighting dans le noyau. Leurs fonctions dans le noyau correspondent :
Ces fonctions répondent à des impératifs cellulaires : en effet, il est indispensable que certains métabolites soient synthétisés dans le noyau.
Les enzymes du métabolisme délocalisées dans le noyau participent donc à un processus [métabolisme - transcription des gènes] qui inclue en particulier les modifications épigénétiques. Ce processus semble optimiser les réponses adaptatives qui relient le stress métabolique à des fonctions cellulaires telles que la prolifération ou la différenciation. De plus en plus de preuves indiquent que les changements de localisation subcellulaire des protéines sont médiés par de courts motifs linéaires. La base de données switches.ELM (EMBL) est un recueil de commutateurs moléculaires ("motif-based molecular switches") validés expérimentalement et reposant sur un motif. Elle permet d'étendre les connaissances sur la manière dont les motifs interviennent dans la "prise de décision coopérative" selon le contexte et participent à la régulation cellulaire. |
6. Identification de protéines moonlighting La caractérisation d'une nouvelle protéine implique de trouver la fonction de cette protéine : cependant, on ne recherche pas systématiquement toutes ses fonctions. Le concours "Evaluation critique de l'annotation fonctionnelle ("Critical Assessment of Functional Annotation", CAFA) est un effort continu et global, mené par la communauté scientifique, qui vise à accroître l'annotation de la fonction des protéines par des moyens informatiques. La plupart des fonctions des protéines moonlighting ont été découvertes par hasard. Il n'existe pas encore de méthode efficace pour les prédire. La mise en évidence expérimentale de protéines moonlighting s'appuie sur des études de mutation conduisant à de nouveaux phénotypes.
De plus en plus de programmes de prédiction des fonctions des protéines multifonctionnelles emploient des méthodes d'apprentissage profond ("deep learning"). Les données d'apprentissage sont, entre autres :
|
| 7. Liens Internet et références bibliographiques |
MoonProt : "A Database for Moonlighting Proteins" moonDB : "A Database of Extreme Multifunctional and Moonlighting Proteins" Multitasking Proteins DataBase (moonlighting proteins database) DextMP : "Deep dive into tEXT for predicting Moonlighting Proteins" |
|
Piatigorsky et al. (1988) "Gene sharing by delta-crystallin and argininosuccinate lyase" Proc. Natl. Acad. Sci. 85, 3479 - 3483 Jeffery C.J. (1999) "Moonlighting proteins" Trends Biochem. Sci. 24, 8 - 11 Hentze & Preiss (2010) "The REM phase of gene regulation" Trends Biochem. Sci.35, 423 - 426 |
|
Mani et al. (2015) "MoonProt: a database for proteins that are known to moonlight" Nucleic Acids Res. 43, D277 - D282 Zhang et al. (2015) "Emerging roles of lysine methylation on non-histone proteins" Cell. Mol. Life Sci. 72, 4257 - 4272 Chapple et al. (2015) "Extreme multifunctional proteins identified from a human protein interaction network" Nat. Commun. 6, 7412 Castello et al. (2015) "Metabolic Enzymes Enjoying New Partnerships as RNA-Binding Proteins" Trends Endocrinol. Metab. 26, 746 - 757 |
|
Boukouris et al. (2016) "Metabolic Enzymes Moonlighting in the Nucleus: Metabolic Regulation of Gene Transcription" Trends Biochem. Sci. 41, 712 - 730 Lu & Hunter (2018) "Metabolic Kinases Moonlighting as Protein Kinases" Trends Biochem. Sci. 43, 301 - 310 Snaebjornsson & Schulze (2018) "Non-canonical functions of enzymes facilitate cross-talk between cell metabolic and regulatory pathways" Exp. Mol. Med. 50, 34 |
|
Gonzalez-Arzola et al. (2019) "New moonlighting functions of mitochondrial cytochrome c in the cytoplasm and nucleus" FEBS Lett. doi: 10.1002/1873-3468 Harvey et al. (2019) "The Diverse Functional Roles of Elongation Factor Tu (EF-Tu) in Microbial Pathogenesis" Front. Microbiol. 10, 2351 |
![]()