
Le cycle de Benson, Calvin & Bassham - la RuBisCO |
| Tweet |
|
|
1. Introduction 2. Première étape du cycle : la ribulose 1,5-bisphosphate carboxylase - oxygénase ou RuBisCO
|
3. Réaction de carboxylation vs. réaction d'oxygénation (la photorespiration) 4. Biosynthèse des sous-unités de la RuBisCO 5. Les autres étapes du cycle RPP 6. Régulation du cycle RPP 7. Application : étude protéomique de la RuBisCO 8. Liens Internet et références bibliographiques |
|
1. Introduction Les réactions photochimiques de la photosynthèse forment de l'ATP et du NADPH + H+ qui vont permettre la réduction du CO2 en glucides (CH2O) par des réactions thermochimiques.
Le cycle de Benson, Calvin & Bassham répond à ces critères. Dans la littérature, ce cycle est dénommé de diverses manières :
Ce cycle est basé sur les travaux de Ruben (datant de 1930) puis ceux de Kamen et Bassham. Melvin Calvin (1911-1997) a reçu le Prix Nobel en 1961 pour ses travaux sur la photosynthèse et l'assimilation du CO2. Voir le montage de la "lollipop", dispositif de Melvin Calvin, Andrew Benson et James Bassham (CalTech - 1950) contenant des chlorelles (Chlorella vulgaris). Cette phase ne nécessite pas de lumière contrairement aux réactions photochimiques de la phase lumineuse. Cependant, chez la plupart des végétaux, le cycle de Calvin se déroule de jour afin que les réactions photochimiques régénèrent l'ATP et le NADPH + H+nécessaires à la synthèse des glucides. |
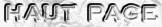
|
2. Première étape du cycle - La ribulose 1,5-bisphosphate carboxylase - oxygénase ou RuBisCO a. Détail de la réaction catalysée par la RuBisCO Le CO2 atteint directement les cellules photosynthétiques par diffusion ou, chez les Plantes supérieures, via des structures de l'épiderme foliaires appelés stomates. L'incorporation du CO2 est la première étape du cycle RPP. Elle est catalysée par la ribulose 1,5-bisphosphate carboxylase - oxygénase ou RuBisCO : 3-phospho D-glycérate carboxy-lyase - E.C.4.1.1.39. La RuBisCO représente environ 50% des protéines totales des feuilles, ce qui en fait l'enzyme la plus abondante de la biosphère. La RuBisCO ne catalyse la fixation du CO2 qu'à l'état activé, c'est-à-dire :
Figure ci-dessous, la réaction d'incorporation du CO2 (substrat) sur un ose phosphorylé à 5 carbones : le ribulose 1,5-bisphosphate (RuBP) .
La concentration des intermédiaires du cycle RPP détermine celle du RuBP. Remarque : dans le cycle RPP complet, ce sont 3 molécules de CO2 qui sont incorporées avant que le plus petit intermédiaire en C3 (le glycéraldéhyde 3-phosphate) ne quitte ce cycle. |
b. Structure oligomèrique de la RuBisCO des Plantes supérieures Chez les Plantes supérieures, la RuBisCO est une protéine oligomérique constituée de 16 sous-unités :
La masse molaire totale est environ 560 kDa pour une stoechiométrie L8S8. Les 8 sous-unités L forment un "coeur" octamérique entouré par 2 "couches" de 4 sous-unités S, chaque couche se situant sur les 2 côtés opposés de la macromolécule. Code : MMDB 6150 - Code : PDB 1RCX - Remarque : on dénombre olusieurs dizaines de structures de RuBisCO dans la PDB. Chez les phototrophes autres que les Plantes supérieures, la structure quaternaire de la RuBisCO est variable. Par exemple, chez la bactérie poupre Rhodospirillum rubrum, c'est un homodimère dont les chaînes polypeptidiques ont une séquence homologue à 30% de celle de la grande sous-unité des Plantes supérieures. |
|
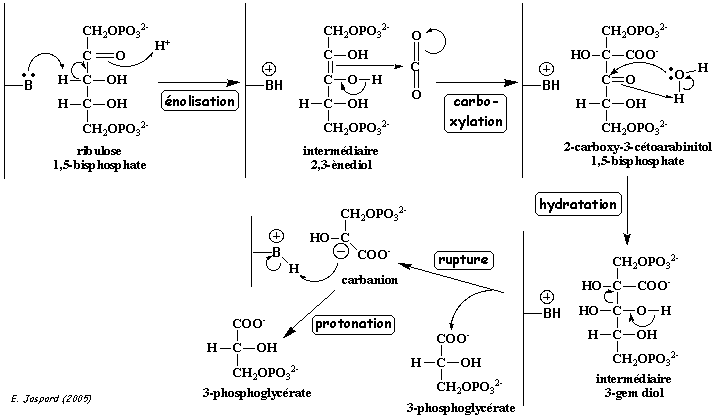
Les 8 sites actifs de la RuBisCO sont organisés autour d'un ion magnésium (en vert dans la figure ci-dessous).
Source : "RuBisCO : molecule of the month" - PDB : 8RUC
L'état activé de la RuBisCO est obtenu quand une une autre molécule de CO2 que celle qui sert de substrat réagit avec la chaîne latérale de la lysine 201 , formant ainsi un carbamate (E-NH-CO2-) stabilisé par Mg2+ (E-NH-CO2-. Mg2+). Le Mg2+ établit aussi une liaison avec le groupe β-COO- de l'aspartate 203.
En conséquence le rôle de cette molécule de CO2 est trés particulier : c'est une molécule activatrice et non pas une molécule substrat de la réaction enzymatique. En effet, elle est :
En conséquence, la nuit, l'ion magnésium peut se fixer à la fois :
|
|
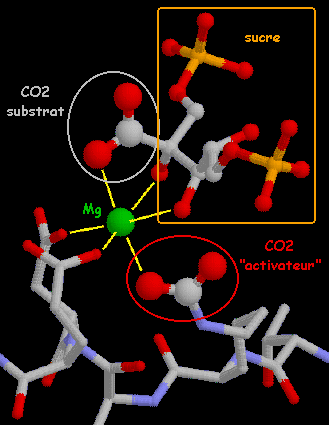
d. Modulation de l'activité de la rubisco par la RuBisCO activase 1. La forme déprotonnée de la lysine 201 (E-NH2) réagit avec le CO2 (activateur) relarguant un proton et formant un carbamate (E-NH-CO2-) stabilisé par Mg2+ (E-NH-CO2-. Mg2+) (figure ci-dessous). 2. La RuBisCO est alors dans sa forme active [encadré bleu de la figure ci-dessous] et fixe le ribulose-1,5-bisphosphate (RuBP) pour former le complexe ternaire (E-NH-CO2- . Mg2+ . RuBP). RuBP est converti en intermédiaire ènediol. Le CO2 ou l'O2 peuvent se fixer au site actif selon que la réaction catalysée est la carboxylation (CO2 substrat), ou l'oxygénation (O2 substrat).
Source : "Rubisco Activase" 3. La forte affinité du ribulose-1,5-bisphosphate pour la forme libre de la RuBisCO (E-NH3+) génère le complexe abortif (E-NH3+ . RuBP) ce qui favorise la formation d'une forme de RuBisCO non-catalytique. 4. Sous un éclairement intense, la RuBisCO activase interagit de manière non covalente avec ce complexe binaire (E-NH3+ . RuBP) et facilite le relarguage du ribulose-1,5-bisphosphate [encadré rouge] libérant ainsi l'enzyme pour le cycle suivant de carbamylation. La RuBisCO activase abaisse le KM pour le CO2 de 12 µM à 2 µM ce qui facilite la réaction de carbamylation. L'importance de ce processus est que la vitesse à laquelle la RuBisCO activase augmente l'activité catalytique de la RuBisCO est liée à l'hydrolyse de l'ATP : 50 molécules d'ATP hydrolysées par site activé de RuBisCO. Il existe 2 isoformes de RuBisCO activase (42 kDa et 46 kDa) qui diffèrent au niveau de leurs extrémités C-terminales du fait d'un épissage alternatif de leur pré-ARN messager commun. |
|
3. Réaction de carboxylation vs. réaction d'oxygénation (la photorespiration) Comme son nom l'indique, la RuBisCO catalyse en fait 2 réactions :
Chez l'algue verte, Chlamydomonas reinhardtii, la boucle entre l'hélice α6 et le feuillet β6 du tonneau α/β du site actif (acides aminés 328 à 342 - PDB : 1GK8) joue un rôle clé dans la discrimination entre les 2 gaz substrats de la RuBisCO, CO2 et O2. Cette boucle adopte une conformation "fermée" ou "ouverte" en présence ou en absence, respectivement, des substrats. Il y a donc compétition entre le CO2 et l'O2 pour le même site de fixation et les vitesses des réactions de carboxylation et d'oxygénation sont conditionnées par les concentrations relatives des deux gaz dans l'atmosphère (tableau ci-dessous). Cette inhibition de la photosynthèse par l'O2 est appelé effet Warburg (1920). |
| composé | concentration dans l'air (%) | kcat(s-1) | KM (µM) |
| CO2 | 0,035 | 3,3 | 9 |
| O2 | 21 | 2,4 | 535 |
| RuBP | ----- | --- | 40 |
| Voir un cours sur les paramètres cinétiques des réactions enzymatiques. | |||
|
Il y a 0,035 % de CO2 dans l'air qui, une fois dans les stomates, n'est plus présent qu'à 0,01 %. Pour un bon fonctionnement de la photosynthèse, il faudrait que la RuBisCO ait une trés forte affinité pour son substrat CO2. Ce n'est pas le cas puisqu'il faudrait que l'air contienne 6% de CO2 pour que la moitié de la vitesse maximale de la réaction de carboxylation soit atteinte. In vivo (c'est-à-dire pour des concentrations : 250 µM O2 et 10 µM CO2), l'activité de carboxylation est cependant 3 à 5 fois celle de l'activité d'oxygénation.
|
|
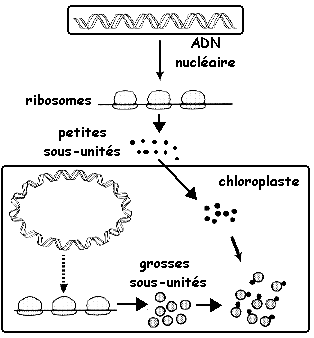
4. Biosynthèse des sous-unités de la RuBisCO
L'assemblage via des molécules chaperonnes a lieu dans le chloroplaste.
Adaptée de : "Physiologie végétale" (1995) - Laval-Martin & Mazliak Cette caractéristique reflète peut-être les origines endosymbiotiques de cet organite. |
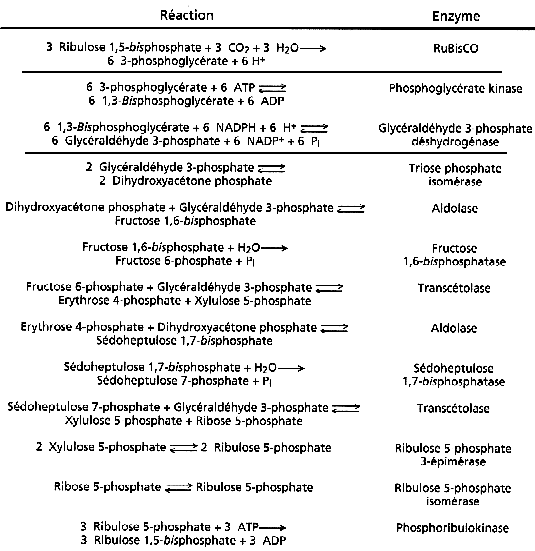
5. Les autres étapes du cycle RPP Le cycle RPP contient 13 réactions.
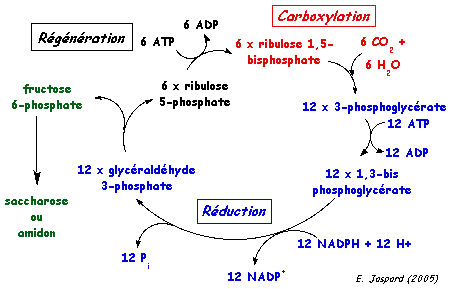
Source : "Principes de Biochimie" Horton et al. (1994) Le cycle RPP peut se décomposer de manière schématique en 3 phases (figure ci-dessous) :
Réduction du 3-phosphoglycérate (APG) Cette réaction synthétise le premier glucide de la photosynthèse en 2 étapes :
L'ATP et le NADPH sont formés lors des phases lumineuses de la photosynthèse. Régénération du RuBP à partir du ribulose 5-phosphate (RuP) Cette régénération utilise les 5/6ème des trioses phosphate synthétisés. Quand le stock de RuBP est seulement renouvelé, 1/6ème au mieux des trioses peut fournir des réserves glucidiques d'origine photosynthétique à la plante. La phosphoribulokinase (EC 2.7.1.19) a un KM de 0,2 mM et 0,1 mM pour le ribulose 5-phosphate (RuP) et l'ATP, respectivement (les concentrations chloroplastiques de ces substrats étant de 0,1 mM et 2 mM, environ). La valeur : ΔG°'(RuP -> RuBP) = - 21,8 kJ. mol-1 indique que la formation du RuBP est fortement favorisée. Bilan du cycle RPP : 6 CO2 + 18 ATP + 12 [NADPH + H+] + 11 H2O -> fructose 6-phosphate + 18 ADP + 12 NADP+ + 17 Pi La réduction de 1 CO2 en glucide nécessite donc 3 ATP et 2 [NADPH + H+] |
|
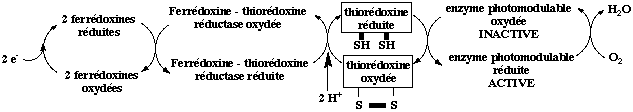
Le contrôle du cycle a lieu aux étapes irréversibles, en particulier les réactions catalysées par : la fructose 1,6-bisphosphatase, la sédoheptulose 1,7-bisphosphatase, la phosphoglycérate kinase, la glycéraldéhyde 3-phosphate déshydrogénase et la phosphoribulokinase. Ces enzymes sont contrôlés par l'intensité lumineuse (sauf la phosphoglycérate kinase), le taux de Mg2+ du stroma et le pH. L'activation par la lumière met en jeu la réduction de ponts disulfure à la surface de ces enzymes, qui modifie leur structure (figure ci-dessous).
Adaptée de : "Principes de Biochimie" Horton et al. (1994)
On notera que les électrons et protons nécessaires à ces réductions sont fournis par la chaîne de transport d'électrons photosynthétique. Ces réductions sont non cycliques et situées du côté oxydant de l'accepteur primaire d'électrons. C'est un exemple de l'utilisation des réactions photochimiques dans un autre but que la synthèse d'ATP et de NADPH. |
| 8. Liens Internet et références bibliographiques |
| "Physiologie végétale" (1995) volume I - "Nutrition et métabolisme", Danielle Laval-Martin et Paul Mazliak, Collection "Méthodes", Hermann / ISBN : 2 7056 6253 7 | |
|
"Etapes du cycle de Calvin" - Université Pierre et Marie Curie (Paris - Jussieu) |
Aller au site |
|
Satagopan
& Spreitzer (2004) "Substitutions at the Asp-473 Latch Residue of Chlamydomonas
Ribulosebisphosphate Carboxylase/Oxygenase Cause Decreases in Carboxylation
Efficiency and CO2/O2 Specificity" J. Biol. Chem. 279, 14240
- 14244
Gerbaud & André (1980) "Effect of CO2, O2, and Light on Photosynthesis and Photorespiration in Wheat" Plant Physiol. 66, 1032 - 1036 |
|
![]()
{kind=link}