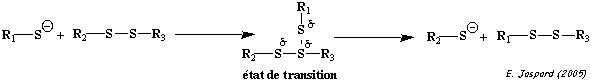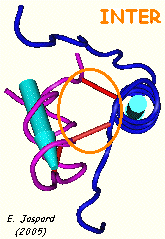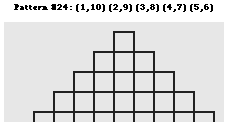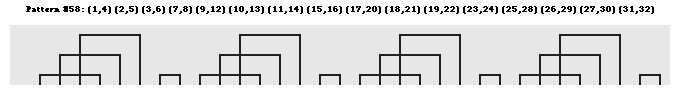| Cystéine et pont disulfure - Prédiction des cystéines impliquées dans un pont disulfure |
| Tweet |
|
|
1ère partie : cystéine et pont disulfure 1. La cystéine 2. Formation d'un pont disulfure
3. Exemples |
4. Fixation des métaux lourds par les métallothionines 2è partie : prédiction des cystéines impliquées dans un pont disulfure 5. Description de la problématique
6. Les différentes approches pour la prédiction de cystéines impliquées dans un pont disulfure 7. Liens Internet et références bibliographiques 8. Module Bioinformatique |
|
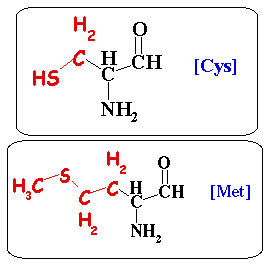
1. La cystéine La cystéine est génétiquement codée par les 2 codons : UGU et UGC. La cystéine est l'acide L-2-amino-3-mercaptopropionique. La chaîne latérale de la cystéine porte un groupement sulfhydryle (-SH) extrêmement réactif. Son pKa est de 8.33. L'autre acide aminé sulfuré, la méthionine, porte un groupe méthyle supplémentaire. La méthionine est plus hydrophobe, d'un encombrement stérique plus important et moins réactive.
|
|
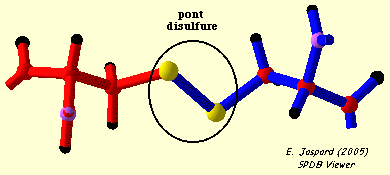
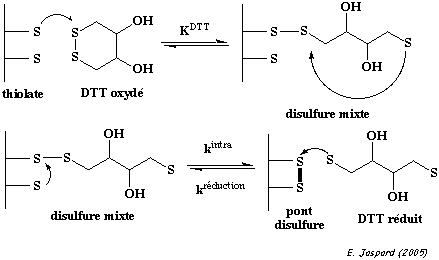
2. Formation d'un pont disulfure Un pont disulfure est une liaison covalente entre 2 atomes de soufre de la chaîne latérale de 2 résidus cystéines (Cys) réduits. Les ponts disulfure sont formés dans un environnement cellulaire oxydant. Le cytoplasme n'étant pas un milieu oxydant, il y a trés peu de protéines intracellulaires qui possèdent des ponts disulfure. Toutes les cystéines d'une protéine ne forment pas un pont disulfure :
Un pont disulfure peut être établi entre 2 cystéines :
Le nombre de ponts disulfure (intra et/ou inter) au sein des protéines varie trés grandement : les toxines, bien qu'étant de petites protéines (20 à 30 acides aminés) peuvent en contenir jusqu'à 5, voire 6. L'énergie libre de liaison qui résulte de la formation d'un pont disulfure est d'environ - 3,5 kcal/mol (voir Czaplewski et al., 2004). C'est une énergie importante qui contribue "sur le papier" de manière trés significative à la stabilisation de la structure tridimentionnelle native de la protéine. Cependant, les ponts disulfure ne jouent pas un rôle déterminant dans le repliement des protéines, puisque les intermédiaires adoptent des conformations transitoires qui peuvent être éloignées de celles qui mettent en jeu ces ponts disulfure. Nombre de ponts susceptibles d'être formés Exemple : quel est le nombre de combinaisons permettant de former 3 ponts disulfures à partir de 6 cystéines ?
Soit : 5 x 3 x 1 = 15 combinaisons. De manière générale : pour n ponts disulfures et 2n cystéines, il existe [(2n)! / 2nn!] combinaisons. Application : script (Python) qui détermine la position des cystéines impliquées dans un pont disulfure sur la base d'un fichier PDB. La distance entre les 2 carbones a de 2 résidus cystéine impliqués dans un pont disulfure est de 4.2 Å (Czaplewski et al., 2004) à 7.5 Å.
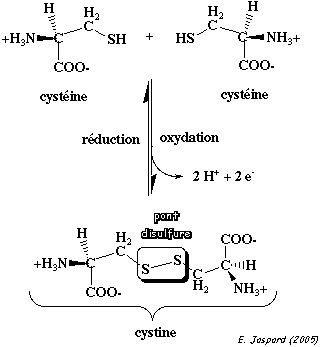
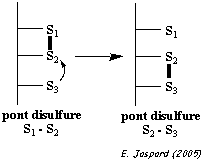
L'entité structurale formé par 2 cystéines reliées par un pont disulfure s'appelle une cystine.
La formation d'un pont disulfure est une réaction d'oxydo-réduction réversible : cette réversibilité dépend du potentiel rédox du milieu et du pH. |
|
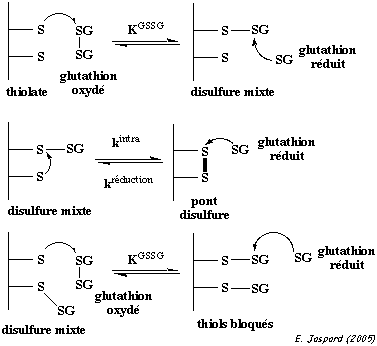
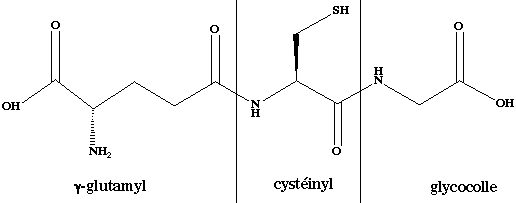
C'est un tripeptide formé de glycine, de cystéine et de glutamate. Cependant, il a une particularité : c'est le groupement carboxylique en position γ de la chaîne latérale du glutamate qui établit la liaison peptidique avec la cystéine. C'est donc le γ-glutamyl-cystéinyl-glycine (structure ci-dessous). Le glutathion existe sous forme réduite ou oxydée. La forme oxydée correspond à l'association de 2 molécules de glutathion reliées par un pont disulfure (G-S-S-G). L'enzyme qui catalyse cette réaction est la glutathion réductase : G-S-S-G + NADPH + H+ <=> 2 G-SH + NADP+
L'une de ses fonctions biologiques est l'élimination de produits dérivés de l'oxygène qui sont hautement réactionnels et donc toxiques pour la cellule :
2 glutathion-SH + H-O-OH -> glutathion-S-S-glutathion + 2 H2O |
|
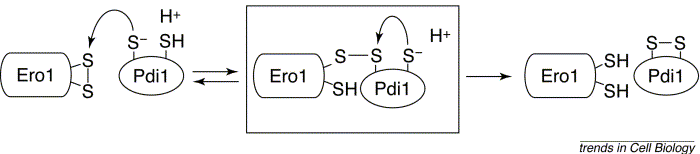
d. Les protéines disulfide isomerases (PDI) et Ero1p Les protéines disulfide isomerases (EC 5.3.4.1) catalysent le ré-arrangement correct dans le réticulum endoplasmique des ponts disulfure des protéines néo-synthétisées. Ce sont des protéines à plusieurs domaines. Pour oxyder les groupements thiols en disulfure des protéines cibles , les cystéines du site actif de la PDI doivent être maintenues dans l'état oxydé. Ero1p est une flavoenzyme associée à la membrane qui forment des ponts disulfure et les transfère à la PDI. Cette dernière peut alors oxyder les cystéines des protéines substrats. Le mécanisme de l'échange [thiol / disulfure] entre Ero1p et la PDI est le suivant :
Source : Frand et al. (2000)
Les 5 ponts disulfure intra-chaînes de Ero1p : CYS 90 - CYS 349, CYS 100 - CYS 105, CYS 143 - CYS 166, CYS 150 - CYS 295, CYS 352 - CYS 355. Ero1p contient :
|
|
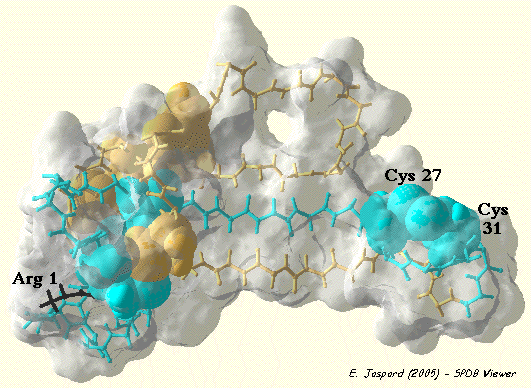
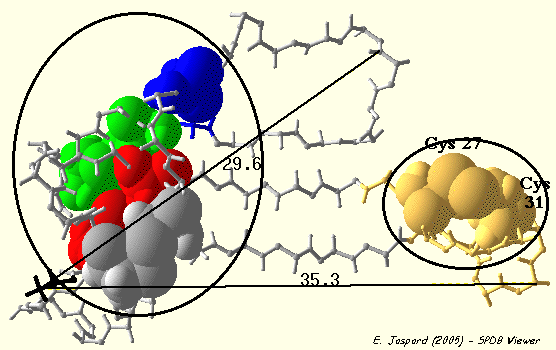
3. Exemples On s'aperçoit que beaucoup de "petites" protéines (selon la classification SCOP - "Structural Classification of Proteins") possèdent des ponts disulfure. La bungarotoxine de venin de serpent est un antagoniste compétitif hautement sélectif et lentement réversible du récepteur nicotinique neuronal α3β2. La bungarotoxine est un homodimère (2 x 66 acides aminés). Chaque sous-unité contient 10 cystéines qui forment 5 ponts disulfure intra-chaîne : R1TC3LISPSSTPQTC14PNGQDIC21FLKAQC27DKFC31SIRGPVIEQGC42VATC46PQFRSNYRSLLC58C59TTDNC64NH66 Les 5 ponts disulfure intra-chaîne de la bungarotoxine : CYS 3 - CYS 21, CYS 14 - CYS 42, CYS 27 - CYS 31, CYS 46 - CYS 58, CYS 59 - CYS 64. La mutagénèse dirigée a permis de montrer que :
Voir une étude bioinformatique de la bungarotoxine (travaux dirigés en ligne). |
|
Visualisation de la bungarotoxine de Bungarus multicinctus obtenue par RMN Code PDB : 2NBT
|
|
En 1982, Rees & Lipscomb (J. Mol. Biol. 160, 475 - 498) ont découvert un nouveau type de repliement en déterminant la structure tridimensionnelle de l'inhibiteur de la carboxypeptidase. Ce type de repliement est le suivant : un pont disulfure pénètre dans un "macrocycle" formé par 2 autres ponts disulfure qui connectent des segments de la chaîne polypeptidique. Une telle structure évoque un "noeud" ou "knot" en anglais. Les membres de cette famille structurales sont ainsi appelés "knottin" ou "inhibitor cystine knots". A ce jour :
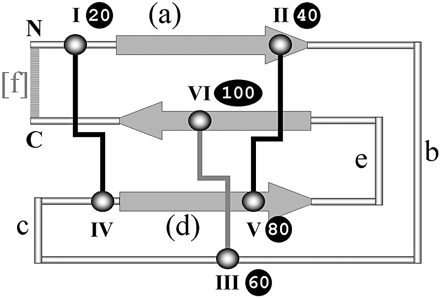
Ci-contre, représentation schématique du repliement des "knottins" :
Source : Gelly et al. (2004) Les flèches indiquent le triple feuillet β antiparallèle trouvé dans la majorité des "knottins". Les cystéines impliquées dans le "noeud" sont marquées par des chiffres romains. Ces 6 cystéines forment 3 ponts disulfure : le pont (III - VI) pénètre dans le "macrocycle" formé par les 2 ponts (I - IV) et (II - V). Les lettres "a" à "f" indiquent la longueur de la boucle entre les cystéines. |
|
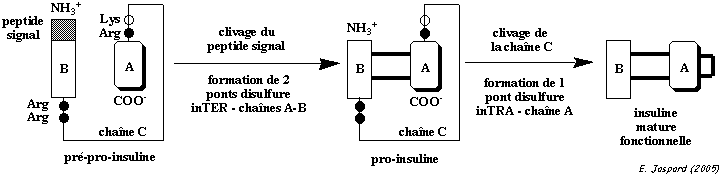
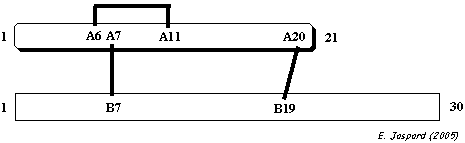
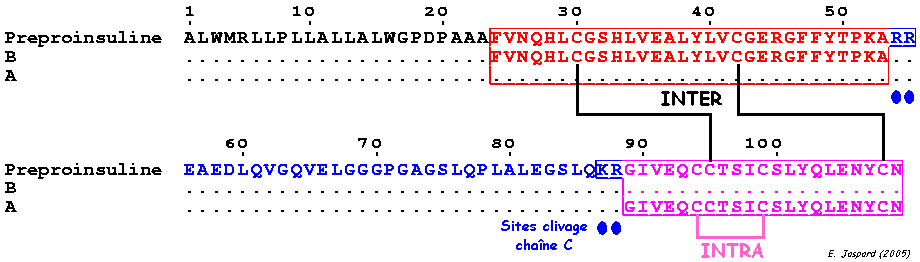
c. Ponts intra-chaîne et inter-chaînes : l'insuline L'insuline est synthétisée sous la forme d'un précurseur, la pré-pro-insuline, dans les cellules β des îlots de Langerhans :
L'insuline adopte alors sa forme fonctionnelle.
Cette maturation par l'élimination du peptide C retarde l'apparition de l'activité hormonale de l'insuline jusqu'à ce qu'elle soit empaquetée dans les granules de sécrétion. |
|
Pont intra-chaîne : CYS 6 chaîne A - CYS 11 chaîne A
Ponts inter-chaînes : CYS 7 chaîne A - CYS 7 chaîne B et CYS 20 chaîne A - CYS 19 chaîne B
Voir un script qui affiche les positions des cystéines impliquées dans un pont disulfure, sur la base d'une structure connue déposée dans la PDB. |
|
d. Ponts intra-chaîne et inter-chaînes : les cathepsines Les cathepsines sont des protéases et il en existe différents types (voir famille des peptidases EC 3.4):
La cathepsine B (EC 3.4.22.1) est une thiol protéase : l'acide aminé du site actif responsable de l'acitivité catalytique est la cystéine 29. L'histidine 199 et une asparagine constituent les 2 autres acides aminés de la triade catalytique. Elle hydrolyse préférentiellement la liaison [Arg-Arg-X] et possède aussi une activité peptidyl-dipeptidase qui libère des dipeptides C-terminaux. La cathepsine B est synthétisée sous la forme d'un précurseur de 339 acides aminés avec un peptide signal (acides aminés 1 à 17). Elles est activée sous la forme d'une chaîne légère (acides aminés 80 à 126 du précurseur) et d'une chaîne lourde (acides aminés 129 à 333). La forme mature est un dimère (1 chaîne légère et 1 chaîne lourde) qui contient 6 ponts disulfure :
|
|
4. Fixation des métaux lourds par les métallothionines Les métallothionines sont des protéines de 60 à 68 acides aminés dont 20 cystéines. Elles constituent une super famille de protéines qui contient 15 sous-familles (animaux, plantes et procaryotes). Elles fixent les métaux lourds tels que le cuivre, le zinc, le cadmium et le mercure. Quand le métal se fixe au groupe sulfhydryle, les ponts disulfure sont rompus et la protéine perd sa structure, la rendant ainsi non fonctionnelle. Les métallothionines contiennent 2 domaines de fixation du métal. Cette fixation se fait via les cystéines de chaque domaine : domaine alpha (11 cystéines - 4 ions fixés) et domaine beta (9 cystéines - 3 ions fixés). Les métallothionines jouent également un rôle protecteur vis-à-vis des espèces oxygénées réactives qui comportent des radicaux libres. L'albumine, la transferrine, la ceruloplasmine, la myoglobine et la ferritine font partie de ce groupe d'antioxidants protéiques. |
|
2ème partie : prédiction des cystéines impliquées dans un pont disulfure |
|
5. Description de la problématique a. Séquence en acides aminés et structure tridimensionnelle de la bungarotoxine R1TC3LISPSSTPQTC14PNGQDIC21FLKAQC27DKFC31SIRGPVIEQGC42VATC46PQFRSNYRSLLC58C59TTDNC64NH66
Oswald et al. (1997) - Code accès : PDB 2NBT
La répartition des cystéines dans la séquence en acides aminés et la répartition spatiale des ponts disulfure ne sont pas corrélés de manière évidente. On remarque 2 cystéines contigües (C58 et C59) impliquées dans 2 ponts distincts. |
|
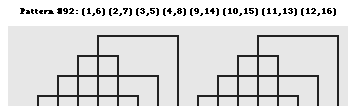
b. Un trés grand nombre de configurations des ponts disulfure Il existe un trés grand nombre nombre d'agencement des ponts disulfure les uns par rapport aux autres au sein des protéines. Lenffer et al. (2004) qui ont développé le programme (et le recueil de données) "CysView" ont recensé 224 configurations, allant de 1 pont (882 séquences) à 36 ponts disulfure (1 séquence). En voici quelques exemples.
Source : "CysView" Le précurseur de la pappalysine 1 du plasma humain (EC 3.4.24.79) contient 36 ponts disulfure (traits gris sur la figure ci-dessous).
Source : PFAM ("Protein families database of alignments and HMMs") |
|
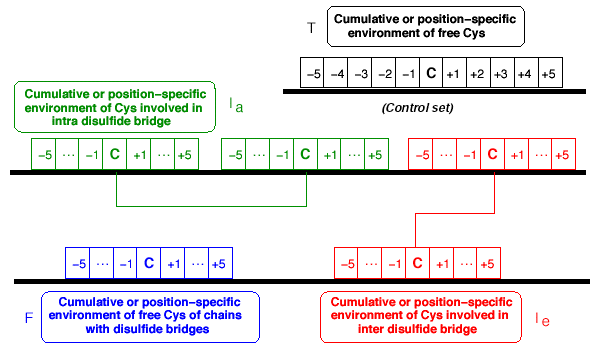
c. Définition d'une fenêtre d'environnement autour des cystéines L'étude de Fariselli et al. (1999) a porté sur une fenêtre de 11 acides aminés autour d'une cystéine (AA-5 ... - C - ... AA-5). Les résultats montrent que la formation d'un pont disulfure par la cystéine centrale C est grandement favorisée si l'on trouve dans cet environnement :
Exemple d'un environnement en acides aminés autour de cystéines impliquées dans des ponts disulfure : signature "Prosite" de la famille des α conotoxines (séquence consensus). C - C - [SHYN] - X(0,1) - [PRG] - [RPATV] - C - [ARMFNHG] - X(0,4) - [QWHDGENFYVP] - [RIVYLGSDW] - C Selon cette notation, on remarque que cet environnement n'est pas défini de manière univoque. En d'autres termes, la même cystéine est impliquée dans un pont disulfure que l'on trouve les acides aminés [R ou P ou A ou T ou V] dans le "voisinage gauche" et que l'on trouve les acides aminés [A ou R ou M ou F ou N ou H ou G] dans le "voisinage droite" . Celà signifie un nombre impressionant de séquences distinctes compatibles avec la formation d'un pont disulfure. Figure ci-dessous, exemple d'un environnement. (J.-M. Richer, G. Hunault & E. Jaspard) :
Celà amène à redéfinir ce voisinage non plus selon l'alphabet à 1 lettre des acides aminés (la séquence primaire), mais selon un nouvel alphabet qui tient compte des propriétés physico - chimiques des acides aminés. |
| 7. Liens Internet et références bibliographiques | |
| Cystéine et pont disulfure | |
|
"Why cysteine is special ?" - Jacek Leluk Frand et al. (2000) "Pathways for protein disulphide bond formation" Trends in Cell Biology 10, 203 - 210 "Metallothionein" : Site du département de Biochimie (Université de Zurich) et "Swiss Institute of Bioinformatics" (SIB) |
|
|
Gross et al. (2004) "Structure of Ero1p, Source of Disulfide Bonds for Oxidative Protein Folding in the Cell" Cell 117, 601 - 610 "The KNOTTIN website and database" : A new information system dedicated to the knottin scaffold (Université de Montpellier - CNRS - INSERM) Gelly et al. (2004) "The KNOTTIN website and database : a new information system dedicated to the knottin scaffold" Nucleic Acids Res. 32, D 156 - 159 |
----- |
| Prédiction des cystéines impliquées dans un pont disulfure | |
|
PDB-select 25 : Selection of a representative set of PDB chains - Most recent 25% list homologs. |
PDB select 25 |
|
Fariselli et al. (1999) "The role of evolutionary information in predicting the disulfide bonding state of cysteines in proteins" Proteins 36, 340 - 346 Martelli et al. (2002) "Prediction of the disulfide-bonding state of cysteines in proteins at 88% accuracy" Protein Science 11, 2735 - 2739 Martelli et al. (2002) "Prediction of the disulfide bonding state of cysteines in proteins with hidden neural networks" Protein Engineering 15, 951 - 953 Mucchielli-Giorgi et al. (2002) "Predicting the disulfide bonding state of cysteines using protein descriptors" Proteins 46, 243 - 249 Czaplewski et al. (2004) "Prediction of the structures of proteins with the UNRES force field, including dynamic formation and breaking of disulfide bonds" Protein Engineering, Design and Selection 17, 29 - 36 Chen et al. (2004) "Prediction of the bonding states of cysteines using the support vector machines based on multiple feature vectors and cysteine state sequences" Proteins 55, 1036 - 1042 Cheng et al. (2005) "Large-Scale Prediction of Disulphide Bridges Using Kernel Methods, Two-Dimensional Recursive Neural Networks, and Weighted Graph Matching", Proteins, (sous presse) |
Fariselli et al. |
|
Sites et logiciels de prédiction liés aux cystéines et aux ponts disulfure
DISpro (SCRATCH protein predictor) : DIpro is a cysteine disulfide bond predictor based on 2D recurrent neural network, support vector machine, graph matching and regression algorithms. GDAP: Genomic disulfide analysis program. DISULFIND : Cysteines Bonding State and Connectivity Predictor. Web interface to a set of Machine Learning programs trained to predict the disulfide bonding state and connectivity pattern of the cysteines in a given amino acid sequence. DSDBASE : Database on disulphide bonds in proteins that provides information on native disulphides and those which are stereochemically possible between pairs of residues in a protein. Triads : A program for identifying Cys-Cys-Trp triad motifs in protein structures. |
|
![]()