| La glycosylation |
| Tweet |
|
|
1. Introduction 2. La O-glycosylation 3. La N-glycosylation 4. Rôle du dolichol phosphate dans la N-glycosylation |
5. Synthèse de l'oligosaccharide lié au dolichol-pyrophosphate et CDG I 6. L'oligosaccharyl transférase 7. Liens Internet et références bibliographiques |
|
1. Introduction La glycosylation a lieu chez tous les Eucaryotes et a été récemment mise en évidence chez les bactéries. La glycosylation est une modification essentiellement post-traductionnelle qui ajoute des glycanes. Par exemple, les immunoglobulines sont glycosylées. Les protéines glycosylées sont destinées à être sécrétées ou à être intégrées à la membrane plasmique. La glycosylation a lieu dans le réticulum endoplasmique. La terminaison des chaînes de glycanes (hydrolyse de la de la cupule de glucides initiaux, branchements, allongement des chaînes) est faite dans l'appareil de Golgi. L'ensemble des glycanes ajoutés aux protéines forme le glycome. Les chaînes polysaccharidiques sont souvent ramifiées et la partie glucidique des protéines glycosylées varie, en poids, de 1 à 50% de la masse de l'ensemble. Les chaînes d'oses sont liées aux protéines par des liaisons O-glycosidiques ou N-glycosidiques selon leur site d'ancrage. Les chaînes liées par des liaisons O-glycosidiques sont plus courtes (1 à 3 résidus d'oses) et plus variables que celles liées par des liaisons N-glycosidiques qui peuvent former des arborescences. La détermination de la structure des glycoprotéines est actuellement l'un des travaux les plus difficiles. La raison en est simple : chaque ose possède plusieurs hydroxyles libres et chacun peut établir une liaison avec un autre ose ou un autre composé. Ainsi, le nombre de polysaccharides qui peut être formé est immense : par exemple, avec seulement trois oses, il existe plusieurs centaines de configurations. Il existe d'autres types de glycosylation :
Enfin, en plus des différents types de glycosylation, les glycanes eux-mêmes peuvent être modifiés. Cela augmente la diversité des glycoprotéines d'un protéome donné. Ces modifications sont :
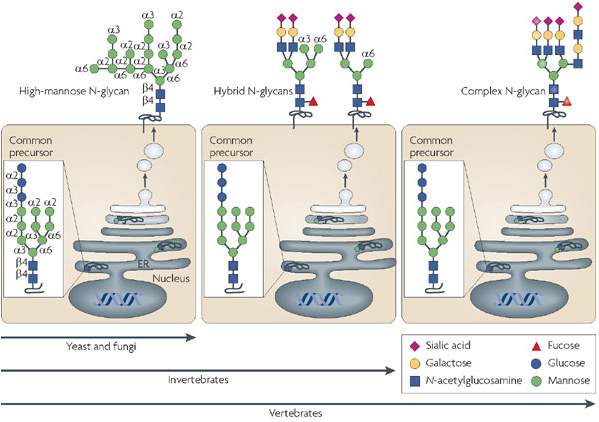
Le degré de complexité des glycanes ajoutés va de paire avec l'évolution de l'organisme considéré. Les principaux saccharides ajoutés dans les chaînes branchées de glycanes sont : N-acétylglucosamine, mannose, fucose, acide N-acétylneuraminique (acide sialique), xylose, galactose, glucose, N-acétylgalactosamine.
Source : Marth & Grewal (2008) |
|
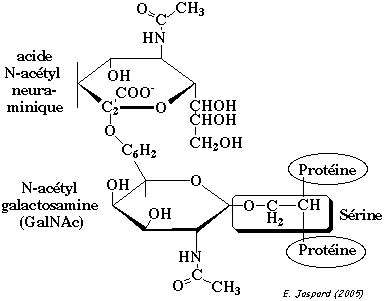
La O-glycosylation est catalysée par des glycoprotéines glycosyltransférases. Cette modification correspond à la liaison établie entre :
Les résidus de sucre sont ajoutés directement sur les acides aminés cibles, les uns après les autres et sous forme activée, c'est-à-dire, liés à un nucléotide. |
|
La N-glycosylation est essentiellement co-traductionnelle : en effet, le glycane est attaché à la chaîne polypeptidique en cours de biosynthèse et pendant son transport dans le réticulum endoplasmique.
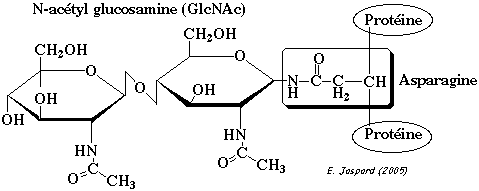
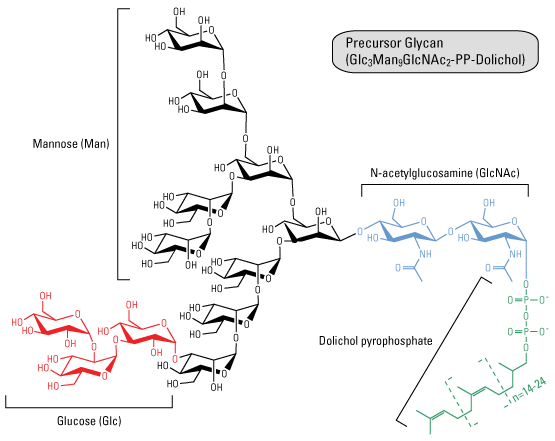
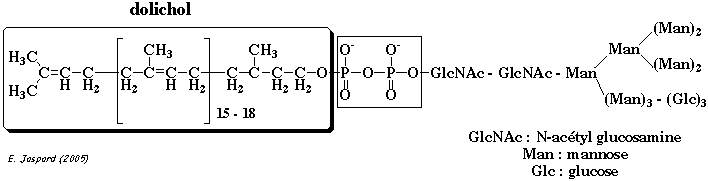
Le N signifie que le glycane est attaché de manière covalente l'azote du groupement amide d'une asparagine (motif Asn-X-[Ser/Thr]). Le sucre immédiatement attachés à l'asparagine est la N-acétylglucosamine. Assemblage des glycanes précurseurs Les oligosaccharides attachés via une liaison N-glycosidique proviennent d'une molécule précurseur contenant 14 oses : la N-acétylglucosamine (GlcNAc), le mannose (Man) et le glucose (Glc).
Source : Pierce
|
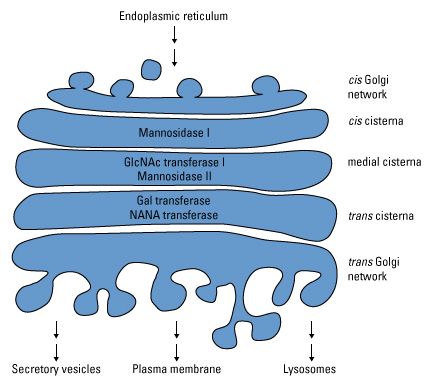
Maturation des glycanes dans l'appareil de Golgi À ce stade de la glycosylation, toutes les glycoprotéines N-glycosylées ont la même structure de glycanes précurseurs. La maturation de ces glycanes dans l'appareil de Golgi combine le clivage et l'addition d'oses pour diversifier les glycanes sur chaque type de glycoprotéines. Comme pour la biosynthèse des glycanes précurseurs, cette voie de maturation pour produire divers oligosaccharides est très ordonnée et chaque étape dépend de l'étape précédente. À cette fin, les enzymes impliquées dans ces modifications sont séparées dans différentes citernes de l'appareil de Golgi.
Source : Pierce Les glycanes destinés à devenir des oligosaccharides complexes sont coupés dans l'appareil de Golgi par la mannosidase I et la mannosidase II et glycosylés par la GlcNAc transférase, résultant en une structure de base commune. Cette structure de base est le substrat de multiples glycosyltransférases qui transfèrent consécutivement des résidus d'oses issus de sucres-nucléotides. Il en résulte des chaînes d'oligosaccharides ramifiées et de longueur variable constituées de GlcNAc, de galactose, d'acide N-acétylneuraminique et de fucose. Toutes les glycoprotéines qui atteignent ce stade sont résistantes à l'hydrolyse de glycanes par l'endo-glycosidase H qui est utilisée expérimentalement pour déterminer si une glycoprotéine contient une forte teneur en mannose ou des oligosaccharides complexes. |
|
Les enzymes de la N-glycosylation
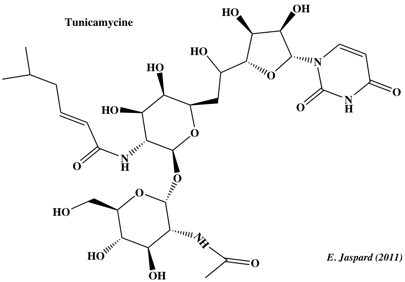
Les molécules de la famille des tunicamycines sont des analogues structuraux de sucre-nucléotide et sont produites par différentes espèces de Streptomyces.
Chez les eucaryotes, elles inhibent l'UDP-N-acétylglucosamine : dolichol phosphate - Glc/NAc-1-P transférase qui catalyse la N-glycosylation initiale de l'asparagine. Chez les bactéries, elles inhibent l'[UDP-N-acétylmuramoyl-pentapeptide : undécaprenol phosphate - Mur/NAc-pentapeptide-1-P transtransférase] qui catalyse une étape précoce de l'assemblage des peptidoglycanes de la paroi cellulaire. |
|
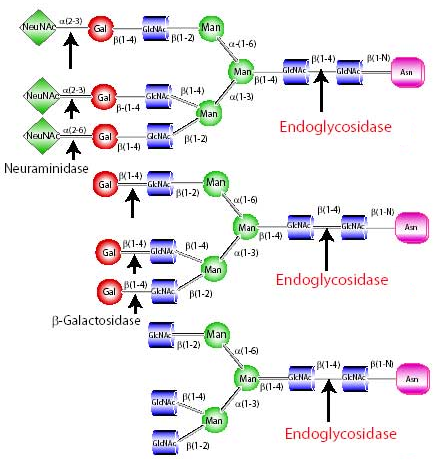
La dé-glycosylation Exemple d'enzymes :
Source : Sigma - Aldrich |
|
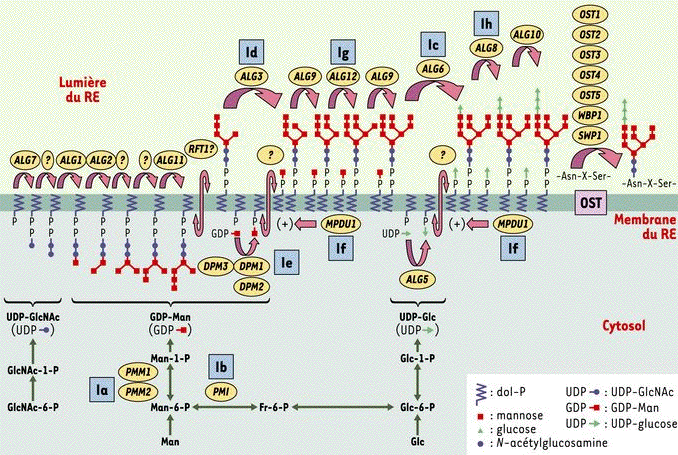
5. Synthèse de l'oligosaccharide lié au dolichol-pyrophosphate et CDG I ("Congenital Disorder of Glycosylation I") - Dupré et al. (2004) Les 7 premières réactions se déroulent sur la face cytosolique de la membrane du réticulum endoplasmique (RE) (partie gauche de la figure ci-dessous). Les monosaccharides sont d'abord activés dans le cytosol en :
Ils sont ensuite greffés au dolichol-phosphate (dol-P).
Source : Dupré et al. (2004) Cercle jaune : nom du gène codant pour l'enzyme responsable de chaque étape. Carré bleu : étapes dont le déficit enzymatique est responsable des CDG (Ia à Ih).
Les réactions suivantes ont lieu dans la lumière du RE et utilisent le dolichol phospho-mannose (dol-P-Man) et le dolichol phospho-glucose (dol-P-Glc) comme donneurs d'oses. L'oligosaccharyl transférase (OST) transfère l'oligosaccharide [Glc3 - Man9 - GlcNAc2] du dolichol pyrophosphate sur une asparagine inclue dans une séquence consensus Asn-X-Ser/Thr d'un polypeptide en cours de biosynthèse dans la lumière du RE. |
|
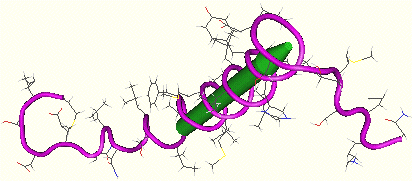
6. L'oligosaccharyl transférase L'oligosaccharyl transférase (EC 2.4.1.119) de la levure est constituée de 9 sous-unités dont 3 (Ost1p, Wbp1p et Stt3p) sont elles-mêmes N-glycosylées. Ci-dessous, la sous-unités Ost4p de l'oligosaccharyl transférase de Saccharomyces cerevisiae (Zubkov et al. - 2003). Code accès : MMDB 26922 - PDB 1RKL
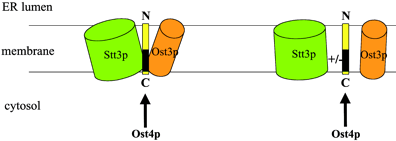
Ost4p est une protéine "minimembranaire" constituée de seulement 36 acides aminés. Ci-dessous, schéma du rôle de la sous-unité Ost4p : elle agirait comme un pont moléculaire médiant les interactions entre les sous-unités Ost3p et Stt3p.
Source : Kim et al. (2003) |
| 7. Liens Internet et références bibliographiques |
|
Dupré et al. (2004) "Les anomalies congénitales de glycosylation des N-glycosylprotéines" Médecine sciences 20 Kim et al. (2003) "Determination of the membrane topology of Ost4p and its subunit interactions in the oligosaccharyltransferase complex in Saccharomyces cerevisiae" PNAS 100, 7460 - 7464 |
|
|
L'ensemble de la voie de biosynthèse des N-glycanes (métabolites et enzymes) "Protein Glycosylation" "Ulysse" - ressources multimedia - Université de Bordeaux |
|
| Marth & Grewal (2008) "Mammalian glycosylation in immunity" Nature Rev. Immunol. 8, 874 - 887 | |
|
"Essentials of Glycobiology" : livre en ligne au NCBI "NetNGlyc 1.0 Server" : logiciel en ligne de prédiction de sites de N-glycosylation |
|
![]()