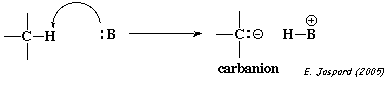| Le lysozyme |
| Tweet |
|
|
1. Généralités 2. Structure du lysozyme 3. Réaction catalysée par le lysozyme |
4. Mécanisme catalytique
5. Liens Internet et références bibliographiques |
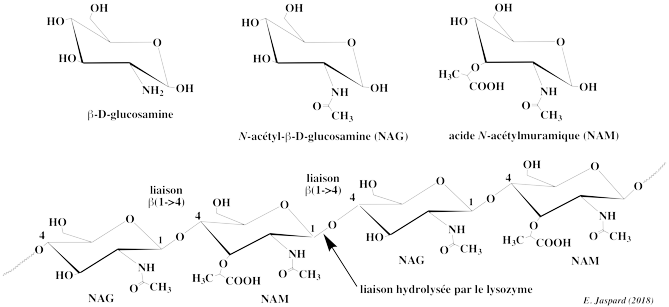
| 1. Généralités Le lysozyme est une enzyme (E.C. 3.2.1.17 - 1,4-β-N-acétylmuramidase C) qui appartient à la famille des O-glycosyl hydrolases. Il hydrolyse la liaison osidique β-(1->4) entre l'acide N-acétylmuramique et la N-acétyl-D-glucosamine des peptidoglycanes et entre les résidus N-acétyl-D-glucosamine des chitodextrines. La première description du lysozyme revient au russe P. Laschtchenko en 1909. Le nom de lysozyme a été donné en 1921 par Alexander Fleming, après qu'il eut observé qu'un "agent des mucus et des larmes" était la cause de la lyse de bactéries. Le lysozyme est trouvé dans de nombreux fluides biologiques, mais la forme la plus étudiée est issue du blanc d'oeuf de poule. Si l'hémoglobine et la myoglobine sont les 2 premières protéines dont on a déterminé la structure 3D, le lysozyme est la première enzyme (David Phillips, 1965). |
|
Pour le seul lysozyme du bactériophage T4, on recense plus de 60 structures cristallographiques de mutants dans la PDB. Le chiffre est encore plus impressionant si l'on comptabilise toutes les structures déterminées pour cette enzyme : près d'un millier. C'est une protéine "globulaire" constituée d'une chaîne polypeptidique de 130 acides aminés (chaîne mature - environ 14,6 kDa) : >1VDQ LYSOZYME C KVFGRCELAAAMKRHGLDNYRGYSLGNWVCAAKFE35SNFNTQATNRNTDGSTD52YGILQINSRWWC NDGRTPGSRNLCNIPCSALLSSDITA SVNCAKKIVSDGNGMNAWVAWRNRCKGTDVQAWIRGCRL129 |
|
Visualisation du lysozyme de Gallus gallus à une résolution de 1,5 Å Code PDB : 1VDQ
|
|
Le lysozyme hydrolyse les polysaccharides. Par exemple, ceux constitutifs de la paroi de certaines bactéries. La paroi bactérienne est constituée par un peptidoglycane, co-polymères de 2 unités d'oses : l'acide N-acétylmuramique (NAM) et la N-acétylglucosamine (NAG), tous deux analogues N-acétylés de la glucosamine. Ces 2 unités d'oses sont reliées par une liaison osidique β-(1->4).
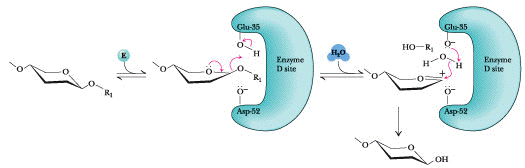
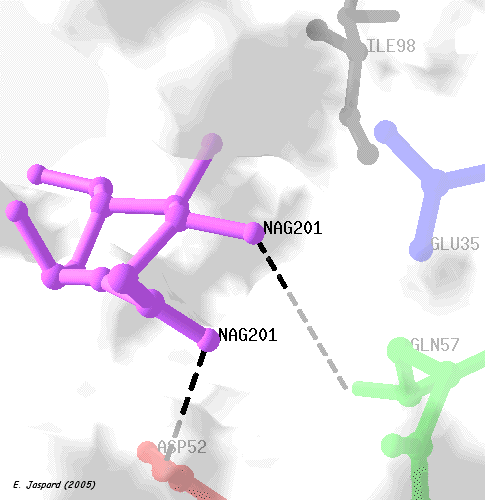
Ce procédé est, par exemple, souvent utilisé lors des premières étapes de la purification de protéines recombinantes sur-exprimées chez Escherichia coli. Dans le même domaine on utilise le plasmide pLysS qui code pour le lysozyme du phage T7, un inhibiteur naturel de l'ARN polymérase de ce phage. La structure tridimensionnelle du lysozyme révèle un site actif qui peut accommoder 6 résidus d'oses du polysaccharide substrat. Chaque résidu d'ose est fixé à un sous-site de fixation et les 6 sous-sites sont dénommés sous-sites A à F.
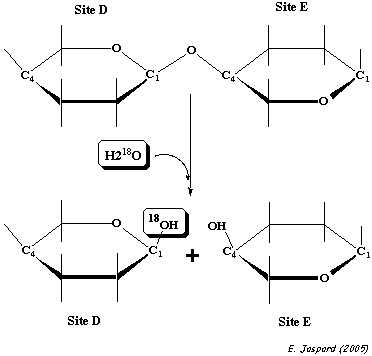
Des expériences utilisant de l'eau enrichie en isotope 18O (H218O) ont montré que l'hydrolyse du polysaccharide substrat libère un ose ayant incorporé18O à la place de l'oxygène lié au C1 fixé au sous-site D : le site d'hydrolyse du substrat est entre les résidus d'oses fixés aux sous-sites D et E. |
4. Mécanisme catalytique a. Rappels sur le mécanisme catalytique acide - base général C'est le mécanisme le plus courant de la catalyse enzymatique. L'accélération de la réaction résulte du transfert d'un proton. Il en existe 2 types (figures ci-dessous) : 1er type : un accepteur de proton (une base, B) peut rompre une liaison C-H en arrachant le proton pour former un carbanion C:-
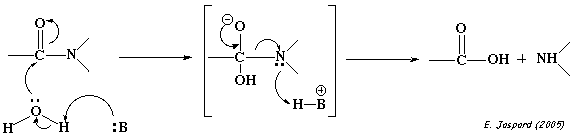
2ème type : un accepteur de proton arrache un proton à une molécule d'eau pour former l'ion OH-. Celui-ci peut se fixer à un carbone qui établit, par exemple, une liaison C-N et cette liaison est rompue.
Bien souvent le proton transféré provient du couple [imidazole / imidazolium] de la chaîne latérale de l'histidine puisqu'elle a un pKa situé entre 6 et 7 et constitue au pH physiologique un groupe [donneur / accepteur] de proton. |
| acide aminé | Glu35 | Asp52 |
| environment | région hydrophobe | région plus polaire |
| état | protonné | ionisé |
| rôle |
1. Glu35 agit d'abord en tant
qu'acide général : donneur de proton à l'oxygène
de la liaison osidique clivée |
Contre-ion négatif du carbocation glycosyle généré au site D lors de l'hydrolyse de la liaison osidique |
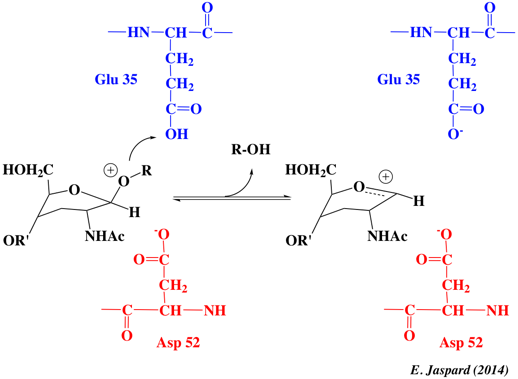
La catalyse implique Glu35 (qui est dans un environnement hydrophobe) et Asp52 (qui est entouré de groupements polaires). Le pKa apparent de Glu35 est donc en principe augmenté : Glu35 est moins susceptible de donner un proton et il acquière une charge négative à pH faible ce qui en fait un meilleur acide général à pH élevés. Des études en fonction du pH démontrent en effet que des chaînes latérales avec un pKa de 3,5 (Asp52) et un pKa de 6,3 (Glu35) sont nécessaires pour la catalyse. |
Mécanisme général Une unité hexa-saccharidique du peptidoglycane est fixée. Il y a distorsion (forme demi-chaise) du NAM du site D (substrat) avant la catalyse. La protonation par Glu35 (pKa élevé) de l'oxygène O sessile du groupement acétal facilite le clivage de la liaison osidique et la stabilisation de la forme résonante d'un ion oxonium positif. Asp52 stabilise l'ion oxonium par interaction électrostatique. La forme demi-chaise déformée du NAM du site D stabilise aussi l'ion oxonium qui exige la coplanarité des substituants attachés au carbone sp2 de la forme résonante du carbocation. Enfin, une molécule d'eau attaque le carbocation, formant l'hémiacétal. Le proton supplémentaire de l'eau est libéré et ce proton est capté par Glu35 (déprotoné) ce qui régénère le système pour une nouvelle catalyse acide général.
La structure tridimentionnelle du complexe [lysozyme-(NAG)3] (un inhibiteur, analogue de substrat) démontre que l'ose du site D adopte effectivement une conformation plane (par opposition à la conformation stable chaise) :
|
|
L'accélération de la réaction catalysée par le lysozyme résulte de :
Source : "Biochemistry - 3rd Edition" (2005) Garrett & Grisham
Figure générée avec Swiss-PDB Viewer (PDB # 1JA7) La constante catalytique du lysozyme est d'environ 0,5 s-1 ce qui est relativement faible par rapport à d'autres enzymes (tableau ci-dessous). Celà signifie que l'hydrolyse de quelques résidus d'oses de la paroi bactérienne suffit : la pression osmotique assure la suite du phénomène de lyse bactérienne. |
| Enzyme | Vitesse non enzymatique (s-1) | Vitesse enzymatique (s-1) | Facteur d'accroissement |
| Chymotrypsine | 4 10-9 | 4 10-2 | 107 |
| Lysozyme | 3 10-9 | 5 10-1 | 2 108 |
| Triose phosphate isomérase | 6 10-7 | 2 103 | 3 109 |
| Fumarase | 2 10-8 | 2 103 | 1011 |
| Uréase | 3 10-10 | 3 104 | 1014 |
| Désaminase d'adénosine | 10-12 | 102 | 1014 |
| Phosphatase alcaline | 10-15 | 102 | 1017 |
| Source : "Principes de Biochimie" Horton et al. (1994), Ed. DeBoecK Universités | |||
| 5. Liens Internet et références bibliographiques |
|
Aibara et al. (2004) "The Crystal Structure Of The Orthorhombic Form Of Hen Egg White Lysozyme At 1.5 Angstroms Resolution" Vocadlo et al. (2001) "Catalysis by hen egg white lysozyme proceeds via a covalent intermediate" Nature, 412, 835 - 3838 |
![]()