
| Régulation de la glycolyse |
| Tweet |
|
|
1. Régulation des étapes irréversibles de la glycolyse
2. Régulation de l'activité de la phosphofructokinase-1 (PFK-1) |
3. Rôle du fructose 2,6-bisphosphate 4. Structure de la [phosphofructokinase-2 / fructose 2,6-bisphosphatase] 5. Régulation de la [PFK-2/FBPase-2] par le glucagon 6. Rôle et structure de l'insuline 7. La charge énergétique adénylique 8. Rôle de GLUT4, de l'insuline et du glucagon dans la régulation de la glycolyse 9. Liens Internet et références bibliographiques |
|
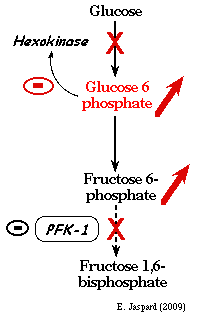
1. Régulation des étapes irréversibles de la glycolyse Ces réactions sont caractérisées par une variation d'énergie libre de Gibbs dans les conditions physiologiques très négatives : elles sont donc thermodynamiquement irréversibles. Ces réactions sont catalysées par des enzymes à régulation allostérique. La modulation de l'activité de ces enzymes est un moyen de contrôler le flux global de la glycolyse. a. Régulation de l'activité de l'hexokinase (E.C. 2.7.1.1) Il en existe 3 isoformes. C'est une enzyme de 102 kDa chez les organismes multicellulaires et environ 50 kDa chez les bactéries.
L'hexokinase est inhibée par l'un des produits de la réaction qu'elle catalyse : le glucose-6-phosphate.
Cette inhibition se fait de deux manières :
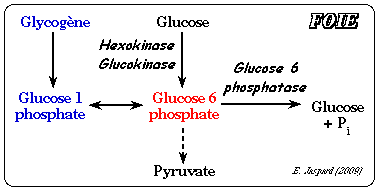
La régulation de l'hexokinase n'est pas le point de contrôle majeur du flux de la glycolyse car une grande partie du glucose-6-phosphate provient de l'hydrolyse du glycogène : glycogène --> glucose 1-phosphate --> isomérisation en glucose 6-phosphate par la phosphoglucomutase. En conséquence la réaction catalysée par l'hexokinase peut être contournée. En revanche, la régulation de l'hexokinase est importante pour réverser la glycolyse et activer la néoglucogénèse quand la concentration en ATP est élevée. |
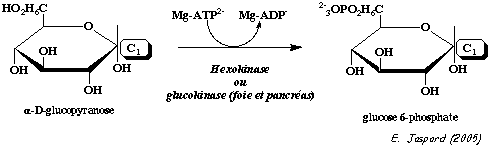
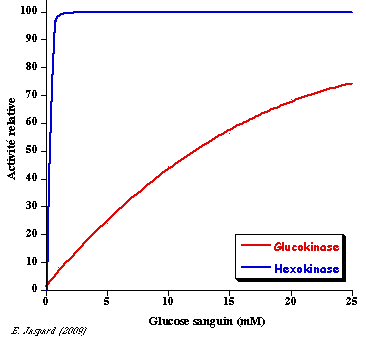
b. Régulation de l'activité de la glucokinase (E.C. 2.7.1.2) C'est l'isoforme IV de l'hexokinase (mammifères) que l'on trouve dans le foie. C'est un monomère de 465 acides aminés d'environ 50 kDa. La glucokinase a un KM élevé pour le glucose et n'est donc active qu'à fortes concentrations de glucose (figure ci-contre) :
Le KM élevé de la glucokinase pour le glucose permet que le foie stocke le glucose sous forme de glycogène quand le taux de glucose sanguin est élevé.
A l'inverse la glucose-6-phosphatase du foie (enzyme de la néoglucogénèse - E.C. 3.1.3.9) catalyse l'hydrolyse du glucose-6-phosphate et le glucose est relargué dans le sang ce qui maintient sa concentration circulante. En conséquence, ces deux enzymes (glucokinase et glucose-6-phosphatase), que l'on ne trouve quasi exclusivement que dans le foie, permettent à celui-ci de contrôler le taux de glucose sanguin.
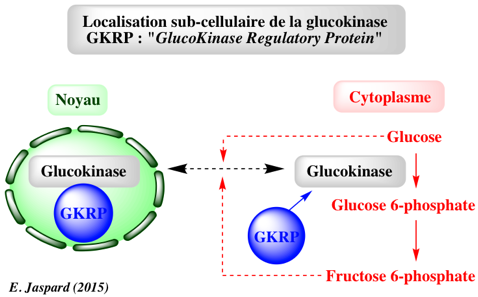
La protéine de régulation de la glucokinase ou GKRP La GKRP ("GlucoKinase Regulatory Protein") est située essentiellement dans le noyau des hépatocytes. La localisation sub-cellulaire de la glucokinase dépend du statut métabolique de la cellule :
La localisation sub-cellulaire de la glucokinase est donc un autre mode de contrôle de son activité et la protéine de régulation GKRP agit comme un senseur métabolique. Voir Shiota et al. (1999). |
|
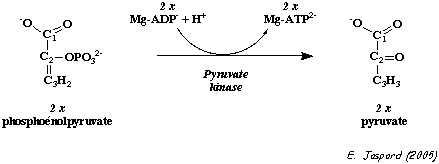
c. Régulation de l'activité de la pyruvate kinase La pyruvate kinase (E.C. 2.7.1.40) est un homotétramère d'une masse de 235 kDa.
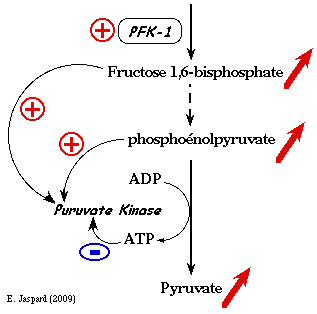
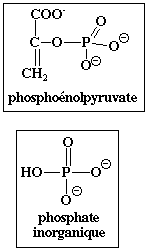
Elle est activée par le fructose 1,6-bisphosphate. Or ce métabolite se situe en amont de la réaction catalysée par la pyruvate kinase : on appelle ce phénomène "feed - forward". Elle est activée par le phosphoénolpyruvate (PEP). Elle est inhibée par l'ATP et par l'alanine (inhibition allostérique).
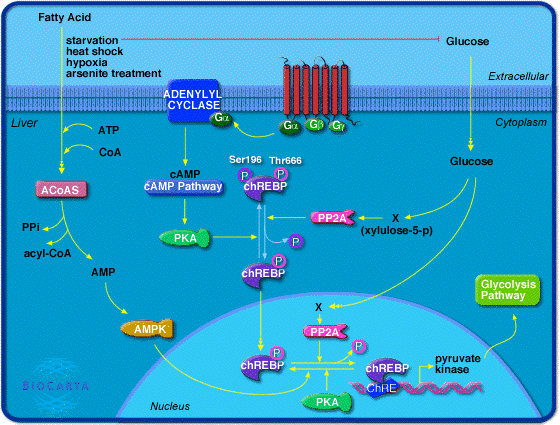
La réaction catalysée par la pyruvate kinase est controlée dans le foie en partie par la modulation de la quantité de pyruvate kinase synthétisée. De fortes concentrations de glucose induisent la délocalisation dans le noyau d'un facteur de transcription (ChREBP) qui active la transcription du gène codant la pyruvate kinase.
Source : "Biocarta" PP2A : protéine phosphatase 2A xylulose-5-phosphate-dépendante ChREBP ("Carbohydrate Responsive Element Binding Protein") est un facteur de transcription de type [hélice-boucle-hélice] "fermeture éclair à leucine" basique ("basic helix-loop-helix/leucine zipper" - bHLH/ZIP) de la famille Mondo. ChREBP est une grosse protéine (96 kDa) multi-domaines :
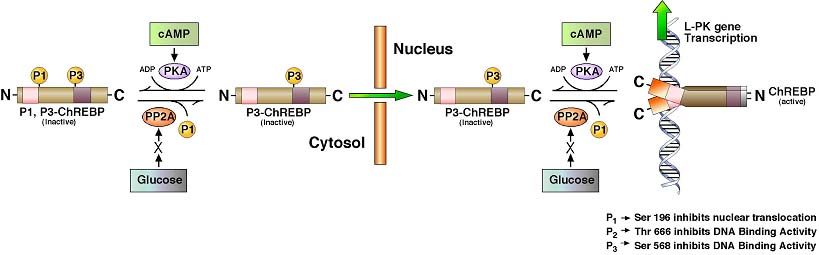
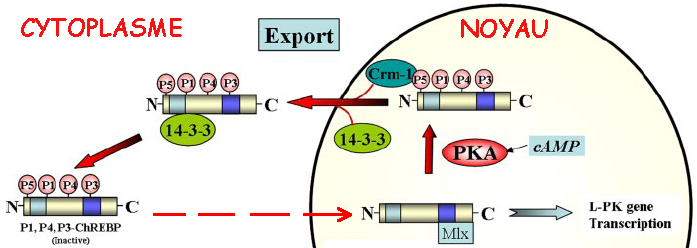
Source : base de données "Reactome" La déphosphorylation (Ser196) de ChREBP par la PP2A (voir "Régulation de la [PFK-2/FBPase-2] par le glucagon"), en réponse à des signaux associés à une nourriture abondante (fortes concentrations de glucose), lui permet d'entrer dans le noyau. ChREBP ainsi activé se fixe sur le motif de l'élément de réponse ChRE (séquence canonique "E box" 5'CACGTG3') de l'ADN près des gènes codant la pyruvate kinase, l'acide gras synthase et l'acétyl-CoA carboxylase. L'excès de glucose est alors converti en pyruvate puis en acétyl-CoA, le principal précurseur de la synthèse des acides gras, qui est la forme de stockage de l'énergie à long terme. La localisation sub-cellulaire de ChREBP dépend aussi de l'interaction d'une hélice (acides aminés 125 à 135) de ChREBP avec une protéine 14-3-3 qui maintient ChREBP dans le cytoplasme.
Source : Sakiyama et al. (2008) L'interaction avec la protéine 14-3-3 est facilitée par la phosphorylation de Ser140 et Ser196 adjacentes à cette hélice. Voir Ge et al. (2012) : structure cristallographique du complexe [ChREBP-14-3-3] à une résolution de 2,41 Å (PDB : 4GNT). |
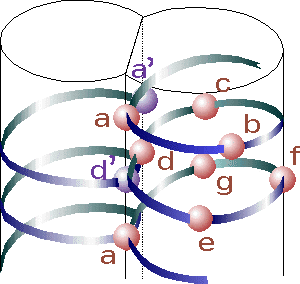
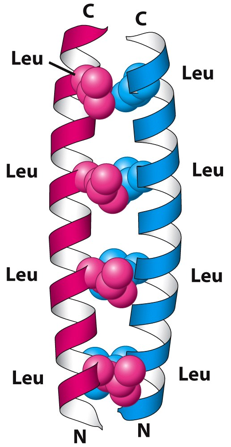
d. Les structures "coiled coil" et le domaine "leucine zipper" de certains facteurs de transcription Une structure dite "coiled coil" (bobine enroulée) est une structure des protéines, où les hélices alpha sont enroulées l'une dans l'autre comme les brins d'une corde tressée. Ces hélices contiennent 3,5 résidus d'acides aminés par tour. Les dimères et les trimères d'hélices sont les types les plus courants. Cette structure a été proposée indépendamment par Linus Pauling et Corey, et par Crick en 1953. Une structure "coiled coil" contient un motif répété de sept acides aminés ("heptad repeat") dont la nomenclature est (abcdefg)n, où a et d sont des acides aminés hydrophobes. Les hélices alpha des structures "coiled coil" sont dites amphipathiques :
Source : "The heptad repeat of the coiled-coil structure"
Un domaine "leucine zipper" est une structure de type "coiled coil" : c'est une structure super-secondaire constituée de deux hélices alpha parallèles enroulées l'une dans l'autre sur la gauche ("left-handed parallel dimeric coiled-coil") et stabilisée par des résidus leucine.
Source : "Oregon state University" Au sein du domaine "leucine zipper", à chaque deuxième tour d'hélice une leucine est en contact direct avec une leucine de l'autre hélice. Le domaine "leucine zipper" est le domaine de dimérisation des facteurs de transcription de la super-classe bZIP ("Basic helix-loop-helix/leucine zipper") des Eukaryotes. Cette super classe contient les classes ou familles suivantes : "Leucine zipper factors", "Helix loop helix factors", "Helix loop helix/leucine zipper factors", "NF family", "RF-X family" et "bHSH family". Les facteurs de transcription CREB ("cAMP response element-binding protein") et GCN4 ("General control protein GCN4") appartiennent également à la super-classe bZIP. |
|
Visualisation du complexe [CREB-CRE] de Mus musculus à une résolution de 3 Å Code PDB : 1DH3 La structure représente les acides aminés 285 à 339 de CREB lié à un désoxyribonucléotides de 21 nucléotides qui englobent l'élément de réponse AMPc de la somatostatine de 8 paires de base.
|
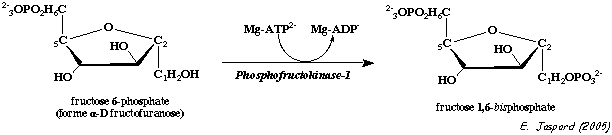
2. Régulation de l'activité de la 6-phosphofructokinase-1 ou PFK-1 (E.C. 2.7.1.11)
La PFK-1 de Escherichia coli est un homotétramère de 320 acides aminés (sous-unité : 34.800 Da). La structure des PFK1 des Eucaryotes est beaucoup plus complexe : la PFK1 de Pichia pastoris est un dodécamère (αβγ)4 (un tétramère de trimères - symétrie D2) d'une masse molaire de environ 106 Da. Ses dimensions sont 161 x 157 x 233 Å. La PFK-1 joue un rôle primordial dans la régulation du flux de la glycolyse. En effet, l'activité de cette enzyme est régulée par la concentration de ces substrats (l'ATP et le fructose 6-phosphate) mais aussi par celles de de nombreux effecteurs liés à la production d'énergie (ATP) par la phosphorylation oxydative. Ces effecteurs sont :
|
|
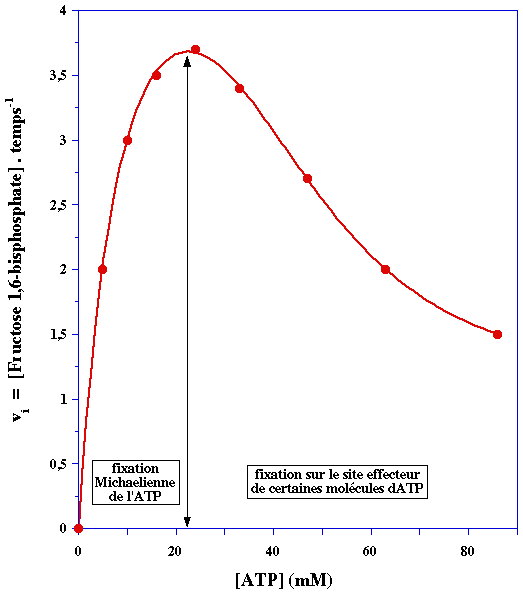
L'ATP est un cas particulier : c'est l'un des deux substrats de la PFK-1 mais c'est aussi un effecteur. En effet la PFK-1 possède :
Partie gauche de la courbe de saturation Elle reflète l'effet de l'ATP sur la vitesse de la réaction enzymatique en tant que substrat. L'allure hyperbolique de cette première partie de la courbe indique que la fixation de l'ATP aux 4 sites catalytiques (la PFK-1 est un homotétramère) s'effectue selon un mécanisme Michaelien (voir le cours). Celà signifie qu'il n'y a pas d'effet coopératif dans la fixation des molécules d'ATP. Partie droite de la courbe de saturation A partir d'une certaine concentration, des molécules d'ATP se fixent sur un site qui n'est pas le site catalytique : ce site est lié à la régulation de l'activité catalytique. C'est un site dit effecteur. Certaines molécules d'ATP n'agissent plus comme substrat mais comme effecteur :
En conséquence, une partie des molécules d'enzyme étant inactive, la concentration réelle de complexe enzyme - substrat ([ES]) est moindre que la concentration d'enzyme. La vitesse de catalyse diminue puisque : vi = kcat x [ES]. |
|
Les différentes phosphofructokinases : nomenclatures et réactions
catalysées Les liens renvoient vers la base de données "Expasy". |
||
| EC | Nomenclature | Réaction catalysée |
| 2.7.1.11 |
|
ATP + D-fructose 6-phosphate ---> ADP + D-fructose 1,6-bisphosphate |
| 2.7.1.56 |
|
ATP + D-fructose 1-phosphate ---> ADP + D-fructose 1,6-bisphosphate |
| 2.7.1.90 |
|
diphosphate + D-fructose 6-phosphate ---> phosphate + D-fructose 1,6-bisphosphate |
| 2.7.1.105 |
|
ATP + D-fructose 6-phosphate <==> ADP + D-fructose 2,6-bisphosphate |
| 2.7.1.146 |
|
ADP + D-fructose 6-phosphate ---> AMP + D-fructose 1,6-bisphosphate |
|
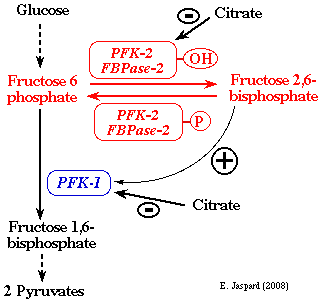
3. Rôle du fructose 2,6-bisphosphate Le fructose 2,6-bisphosphate ou F2,6BP a été découvert en 1980 (Van Schaftingen, Hue & Hers (1980) Biochem. J. 192, 897 - 991). On le trouve chez tous les organismes à l'exception des bactéries.
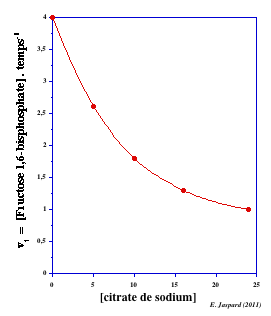
Le F2,6BP est un puissant activateur de la PFK-1 (sauf chez certains protistes pour lesquels il est activateur de la pyruvate kinase).
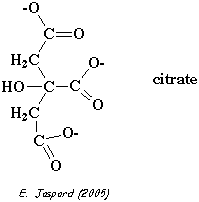
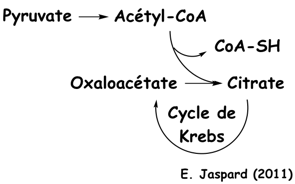
Dans le foie, le F2,6BP est un inhibiteur de la fructose-1,6-bisphosphatase, une enzyme de la néoglucogénèse. Le citrate est un intermédiaire du cycle de Krebs.
Or comme le F2,6BP est un activateur de la PFK-1, celà amplifie l'effet inhibiteur de la PFK-1 par le citrate.
|
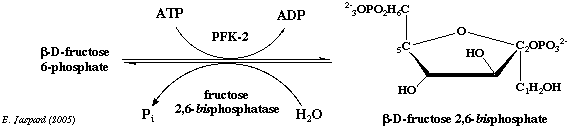
4. Structure de la [phosphofructokinase-2 / fructose 2,6-bisphosphatase] La [PFK-2/FBPase-2] fonctionne sous la forme d'un homodimère. La [PFK-2/FBPase-2] est donc une enzyme bi-fonctionnelle puisque chaque monomère contient 2 domaines ayant chacun une activité enzymatique distincte. Par exemple, l'isoforme du coeur de la [PFK-2/FBPase-2] est une enzyme de 505 acides aminés chez l'Homme (Uniprot : O60825) :
|
|
Visualisation de la [PFK-2/FBPase-2] de l'Homme à une résolution de 2,1 Å Code PDB : 2AXN |
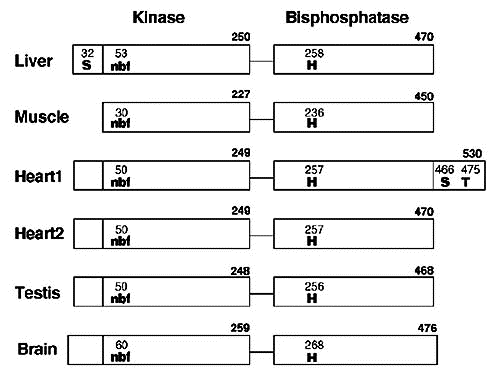
Il existe plusieurs isoenzymes de la [PFK-2/FBPase-2] chez les mammifères. Ci-dessous, représentation des domaines de différentes [PFK-2/FBPase-2] :
Source : Okar et al. (2001)
|
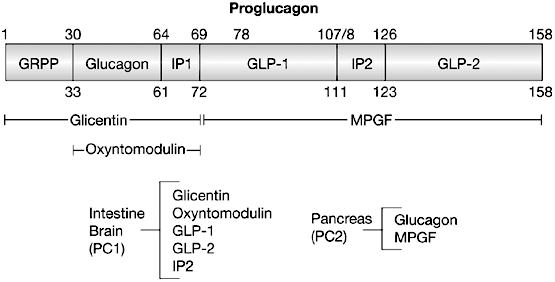
5. Régulation de la [PFK-2/FBPase-2] par le glucagon Le glucagon est une hormone produite par le pancréas quand le taux de glucose sanguin baisse. Le glucagon est synthétisé sous forme d'un précurseur : le proglucagon. Le proglucagon est hydrolysé en 8 chaînes polypeptidiques :
Source : Drucker (2005) Nat. Clin. Prac. Endocrin. Metabol.
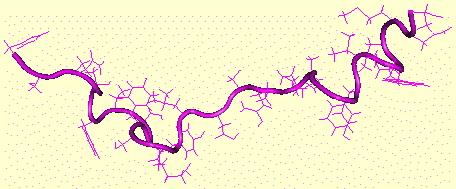
GLP-1 ("glucagon-like peptide 1") est une hormone peptidique de 30 acides aminés. Il potentialise la sécrétion d'insuline stimulée par le glucose et augmente la biosynthèse d'insuline via l'induction de la transcription du gène codant l'insuline. GLP-1 stimule l'adénylate cyclase et la phospholipase C avec une activation subséquente de la protéine kinase A et de la protéine kinase C, ce qui conduit à une augmentation du taux de calcium cytosolique dans les cellules pancréatiques et non pancréatiques. GLP-2 est un peptide de 33 acides aminés co-sécrété avec GLP-1, l'oxyntomoduline et la glicentine, par les cellules L entéroendocrines. L'oxyntomoduline est un peptide de 37 acides aminés qui stimule la sécrétion d'insuline, ralentit la vidange gastrique, inhibe la sécrétion d'acide gastrique, stimule l'absorption intestinale du glucose et diminue la sécrétion des enzymes pancréatiques chez le rat. Le glucagon est un peptide de 29 acides aminés.
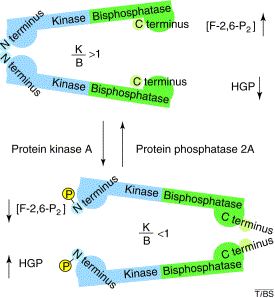
Figure ci-dessus : structure du glucagon de Bos taurus - (Code PDB 1KX6) - Braun et al. (2002). Dans le foie, la PFK-2 est sous le contrôle du glucagon. Quand la concentration du glucagon augmente, la protéine kinase AMP cyclique dépendante (PKA) phosphoryle une sérine ou une thréonine de la [PFK-2/FBPase-2]. Exemples : foie de rat : Ser 32 - coeur de boeuf : Ser 84 - levure : Thr 157. La déphosphorylation est catalysée par la protéine phosphatase 2A xylulose-5-phosphate-dépendante qui est activée par le glucose.
K/B : rapport des activités [PFK-2 / FBPase-2] - HGP : production du glucose par le foie Source : Okar et al. (2001) La phosphorylation :
Il en résulte une baisse de concentration du F2,6BP et la PFK-1 est moins activée. En conséquence, la glycolyse est ralentie quand la concentration du glucagon augmente après une baisse du taux de glucose sanguin. |
|
6. Rôle et structure de l'insuline Le glucose pénètre dans le muscle et le tissu adipeux par l'intermédiaire de transporteurs contrôlés par l'insuline. Une carence en insuline (diabète insulino dépendant) a donc pour conséquence une élévation de la concentration du glucose sanguin.
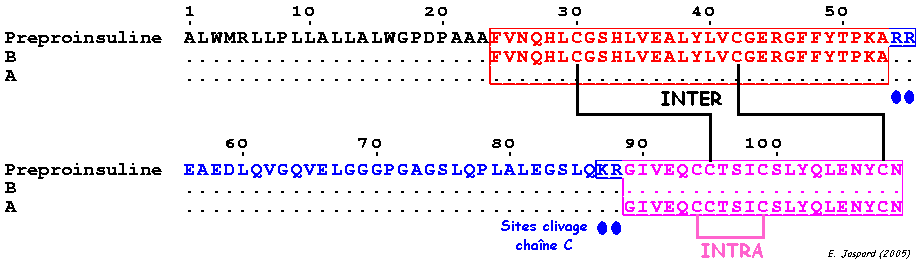
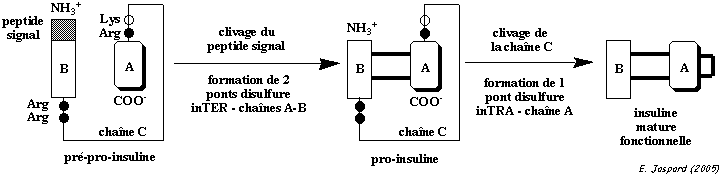
L'insuline est synthétisée sous la forme d'un précurseur, la pré-pro-insuline, dans les cellules β des îlots de Langerhans. Le peptide signal est clivé dans le réticulum endoplasmique.
Cette maturation par l'élimination du peptide C retarde l'apparition de l'activité hormonale de l'insuline jusqu'à ce qu'elle soit empaquetée dans les granules de sécrétion. L'insuline mature est finalement constituée de 4 chaînes polypeptidiques :
Les chaînes [A + B] forment un monomère (figure ci-dessus) et les chaînes [A' +B'] forment un autre monomère. Ces chaînes sont liées pas des ponts disulfure intra- et inter-chaînes :
Voir un développement sur le rôle des transporteurs d'oses (notamment GLUT), de l'insuline et du glucagon dans la régulation de la glycolyse |
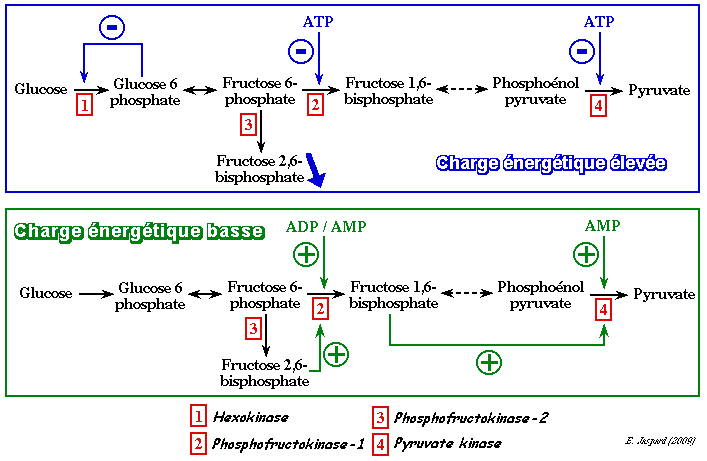
7. La charge énergétique adénylique La charge énergétique adénylique (CEA) de la cellule a été définie par Daniel Atkinson et Gordon Walton (1967) par la relation : [ATP]
+ 1/2 [ADP] Cette relation exprime la fraction molaire en ATP (qui contient 2 liaisons phosphoanhydride à haut potentiel énergétique) plus la moitié de la fraction molaire en ADP (qui n'en contient qu'une). La CEA est donc une mesure de l'énergie disponible à un instant donné dans une cellule, un tissu ou un organisme. Il s'agit d'un paramètre de valeur universelle atteignant des valeurs comparables chez tous les organismes vivants. La CEA est très sensible aux variations de l'environnement interne des organismes (stress) ou de l'environnement extérieur : plus un organisme subit l'effet d'un stress, plus il consomme d'énergie (donc d'ATP) pour contre-balancer ces effets, plus la CEA de cet organisme baisse. En théorie, la valeur de la CEA peut varier de 0 (il n'y a que de l'AMP) à 1 (il n'y a que de l'ATP). Cependant, dans la plupart des cellules, la CEA est trés finement régulée et on observe des variations de 0,7 (forte hydrolyse de l'ATP et de l'ADP en AMP) à 0,95 (typiquement une cellule au repos).
Quand la valeur de la CEA est élevée, l'énergie que contient la cellule est suffisante : la glycolyse est ralentie. Quand la valeur est basse, la cellule a besoin d'énergie : la glycolyse est activée. |
| Résumé des activateurs (A) et des inhibiteurs (I) des enzymes clé de la glycolyse | |||
| hexokinase* | phosphofructokinase 1 | pyruvate kinase | |
| glucose 6-phosphate | I | ||
| fructose 1,6-bisphosphate | A | ||
| fructose 2,6-bisphosphate | A | A (pour certains organismes tels que Kinetoplastida - Trypanosoma) | |
| ATP | I (site
effecteur - effet hétérotrope) |
I | |
| ADP | A | A (substrat - effet homotrope) | |
| AMP | A | I | |
| acétyl CoA | I | ||
| NADH | A puis I | ||
| citrate | I | I | |
| PEP | I | ||
| Pi | A | ||
| insuline | stimulation | ||
| glucagon | inhibition | ||
|
*La glucokinase n'est pas inhibée par le glucose 6-phosphate. La glucokinase possède un motif de fixation de la [PFK-2 / FBPase-2] qui la stabilise dans sa forme active. |
|||
| 9. Liens Internet et références bibliographiques |
|
"Gluconeogenesis" : un superbe site pédagogique pour la biochimie. Voir une excellente animation du flux de la glycolyse (M. Lalanne - Université Laval - Québec) |
|
|
Atkinson & Walton (1967) "Adenosine triphosphate conservation in metabolic regulation. Rat liver citrate cleavage enzyme" J. Biol. Chem. 242, 3239 - 3241 Shiota et al. (1999) "Nuclear Import of Hepatic Glucokinase Depends upon Glucokinase Regulatory Protein, whereas Export Is Due to a Nuclear Export Signal Sequence in Glucokinase" J. Biol. Chem. 274, 37125 - 37130 |
|
|
Okar et al. (2001) "PFK-2/FBPase-2: maker and breaker of the essential biofactor fructose-2,6-bisphosphate" TIBS 26, 30 - 35 Baltrusch & Tiedge (2006) "Glucokinase Regulatory Network in Pancreatic β-Cells and Liver" Diabetes 55, S55 - S64 Sakiyama et al. (2008) "Regulation of Nuclear Import/Export of Carbohydrate Response Element-binding Protein (ChREBP)" J. Biol. Chem. 283, 24899 - 24908 |
|
|
Rider et al. (2004) "6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase: head-to-head with a bifunctional enzyme that controls glycolysis" Biochem. J. 381, 561 - 579 Strater et al. (2011) "Molecular architecture and structural basis of allosteric regulation of eukaryotic phosphofructokinases" FASEB J. 25, 89 - 98 |
![]()
{kind=link}