| Régulation de la dégradation du glycogène |
| Tweet |
|
|
1. Rappels sur le glycogène 2. Cascades de réactions amenant à la dégradation du glycogène 3. Activation des protéines kinases A par l'AMP cyclique (AMPc) 4. La phosphorylase kinase |
5. Régulation par l'inositol 1,4,5-triphosphate (IP3), le 1,2-diacylglycérol (DAG) et le calcium 6. Action de l'adrénaline (dite aussi épinéphrine) 7. Liens Internet et références bibliographiques |
|
1. Rappels sur le glycogène La régulation du métabolisme du glycogène est le prototype de la régulation des sytèmes biochimiques. Les enzymes qui contrôlent ce métabolisme sont aussi celles qui contrôlent d'autres voies métaboliques comme la glycolyse, la synthèse des acides gras, la gluconéogénèse et la synthèse du cholestérol. Le stock intracellulaire de glycogène est utilisé :
L'α-D-glucose est l'ose principalement utilisé dans la glycolyse et il provient de réserves sous forme :
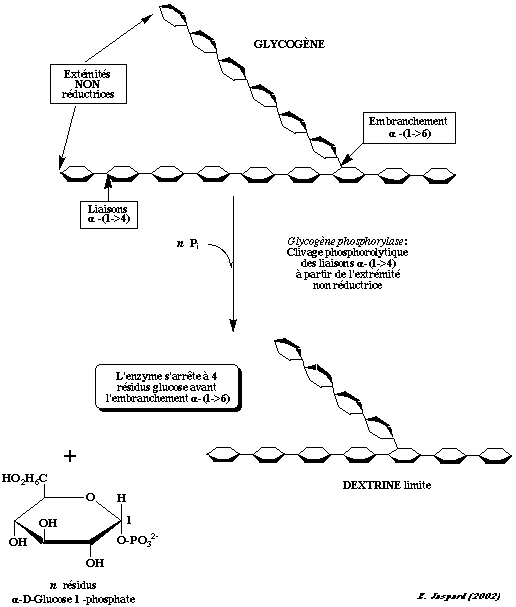
Le glycogène est constitué :
Les résidus glucose du glycogène sont clivés par la glycogène phosphorylase à partir de l'extrémité non réductrice du glycogène. Il s'agit d'une coupure phosphorolytique avec un phosphate inorganique : (α-1,4 chaîne de glycogène)n + Pi <===> (α-1,4 chaîne de glycogène)n-1 + α-D-glucose-1-phosphate
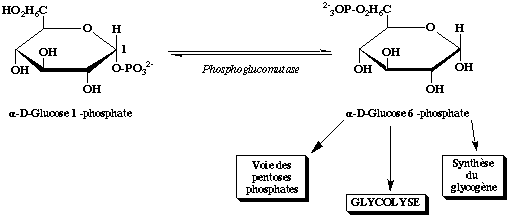
Puis le glucose 1-phosphate formé est transformé en glucose 6-phosphate par la phosphoglucomutase (E.C.5.4.2.2).
Cette enzyme catalyse la réaction dans les deux sens :
|
| Enzyme | Rôle | Caractéristiques |
| glycogène phosphorylase | Catalyse la réaction de l'étape limitante de la dégradation du glycogène | EC
2.4.1.1 / PDB : 1Z8D
/ Environ 5 105 Da / Coenzyme :
pyridoxal Dans le cytoplasme des cellules du foie : 2 sous-unités identiques / muscle : 4 sous-unités |
| glycogène synthase | Catalyse la réaction de l'étape limitante de la synthèse du glycogène | EC 2.4.1.11 / PDB : 2BIS |
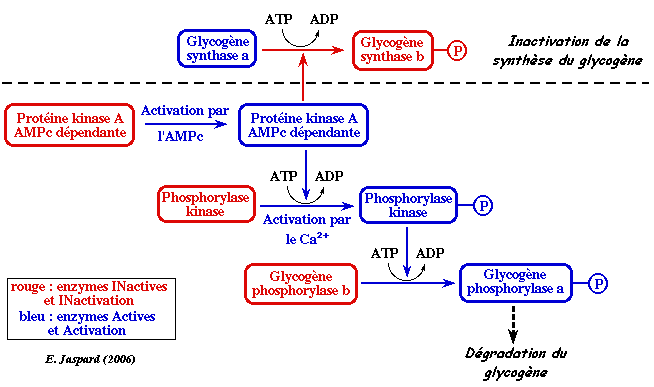
| Ce sont 2 enzymes à formes interconvertibles, lesquelles sont contrôlées par une phosphorylation ou une déphosphorylation de résidus sérine. | active | inactive |
| glycogène phosphorylase | phosphorylée sur la sérine 14 | -OH |
| glycogène synthase | -OH | phosphorylée |
|
complexe le calcium libre dans le cytosol protomère intégré à la phosphorylase kinase |
||
| protéine phosphatase 1 | Catalyse la déphosphorylation des phosphoprotéines du métabolisme du glycogène | |
| inhibiteur-1 | Inhibe la protéine phosphatase 1 | |
Des hormones telles que l'épinephrine (aussi appelée adrénaline), l'insuline et le glucagon régulent l'activité de la glycogène phosphorylase via des voies de signalisation impliquant des protéines G. L'activité de la glycogène phosphorylase b est également contrôlée par des effecteurs allostériques :
La glycogène phosphorylase est la première enzyme à régulation allostérique qui ait été découverte, par Gerty Cori, Carl Cori et Arda Green en 1943. Ils ont démontré que cette enzyme existe sous deux formes (a et b) selon son état phosphorylé. Gerty Cori et Carl Cori ont eu le Prix Nobel en 1947 pour leur description du métabolisme du glycogène. Cori & Green (1943) "Crystalline muscle phosphorylase II prosthetic group" JBC 151, 21–29 |
|
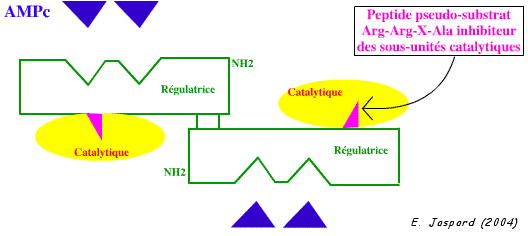
3. Activation des protéines kinases A par l'AMPc Les protéines kinases A sont constituées de 2 sous-unités catalytiques et de 2 sous-unités régulatrices. Les sous-unités catalytiques phosphorylent les résidus Ser ou Thr présents dans la séquence Arg-Arg-X-Ser/Thr. L'inhibition des sous-unités catalytiques par les sous-unités régulatrices résulte de l'occupation du site catalytique par un peptide pseudo-substrat Arg-Arg-X-Ala, qui ne peut être phosphorylé (il n'y a pas de groupement hydroxyle sur la chaîne latérale de l'Ala).
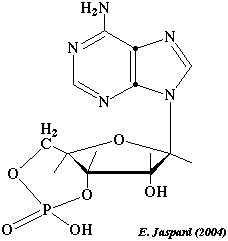
La fixation d'AMPc (figure ci-dessous) sur les 2 sites de chaque sous-unité régulatrice des protéines kinases A induit un changement de conformation de ces sous-unités. Les deux sous-unités catalytiques sont alors relarguées et deviennent actives.
|
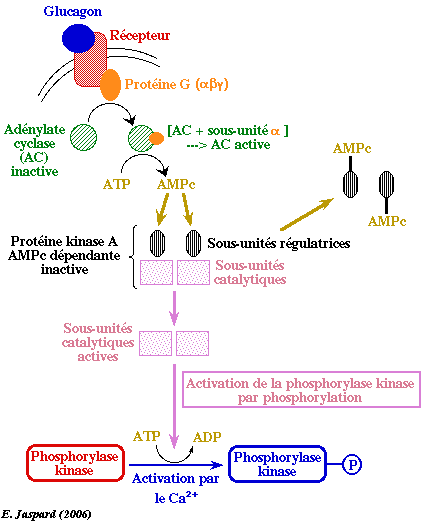
Action du glucagon Le glucagon est une hormone peptidique de 29 acides aminés sécrétée par le pancréas : elle stimule la lipolyse et la conversion des acides gras libres en cétones et inhibe la synthèse et favorise la dégradation des protéines.
Les seules cellules qui possèdent de nombreux récepteurs du glucagon sont celles du foie. Le glucagon agit donc de façon extrêmement sélective. Le glucagon se lie à un récepteur de glucagon qui active une protéine G appelée GS, constituée de 3 sous-unités. La sous-unité α se fixe à l'adénylate cyclase qui est activée : elle convertit l'ATP en AMPc. L'AMPc se fixe aux sous-unités régulatrices de la protéine kinase A ce qui induit la dissociation des sous-unités catalytiques qui phosphorylent la phosphorylase kinase. Cette dernière active à son tour la glycogène phosphorylase en la phosphorylant. Remarque : la phosphorylase kinase active un grand nombre d'autres protéines. Voir l'action du glucagon sur la régulation de la glycolyse. |
|
4. La phosphorylase kinase - EC 2.7.11.19 (EC 2.7.1.38) Première sérine-thréonine protéine kinase découverte en 1959 par Edwin Krebs et al. La phosphorylase kinase est composée de 4 tétramères (16 sous-unités, héxadécamère), chaque tétramère étant constitué de 4 sous-unités distinctes : (αβγδ)4. Sa masse molaire est d'environ 1,3 106 Da.
|
|
Visualisation de la phosphorylase kinase de Oryctolagus cuniculus, à une résolution de 2,4 Å Code PDB : 1QL6 Remarque : sous-unité γ - acides aminés 11 à 291.
|
|
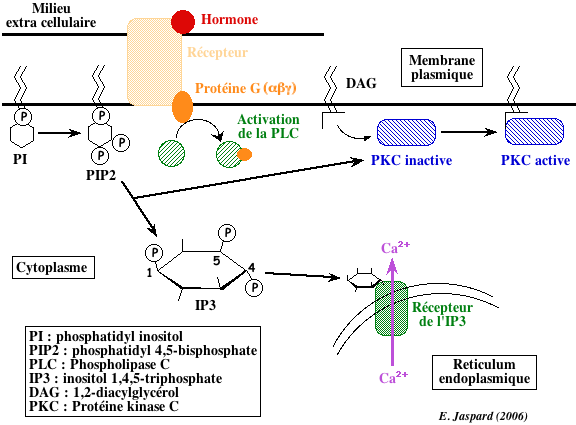
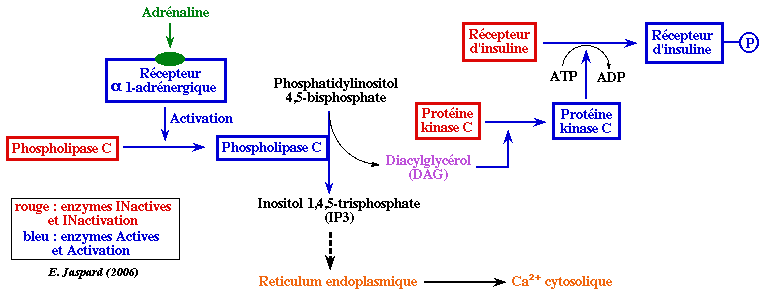
5. Régulation par l'inositol 1,4,5-triphosphate (IP3), le diacylglycérol (DAG) et le calcium L'hydrolyse d'un phospholipide de la membrane plasmique, le phosphatidyl 4,5-biphosphate (PIP2) par la phospholipase C (une enzyme ancrée à la membrane) forme 2 messagers secondaires (secondaires car il transmettent le signal émis par un messager primaire, l'hormone) :
La fonction principale de l'IP3 est la mobilisation du calcium intracellulaire essentiellement stocké dans le réticulum endoplasmique (RE). Il existe des récepteurs de l'IP3 sur la face cytoplasmique de la membrane du RE. Ces récepteurs sont des homotétramères qui forment un canal qui laisse sortir le Ca2+ dans le cytosol lorsque chaque monomère fixe une molécule d'IP3. Le Ca2+ cytosolique se fixe à la calmoduline (voir plus haut).
Le DAG provoque :
La protéine kinase C est activée par la formation d'un complexe quaternaire [protéine kinase C - DAG - phospholipide - Ca2+]. Le phospholipide le plus efficace pour cette activation est la phosphatidyl-sérine. Après activation, la protéine kinase C est transferrée du cytoplasme vers la membrane où elle phosphoryle des protéines sur les résidus sérine et thréonine. |
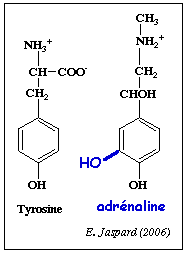
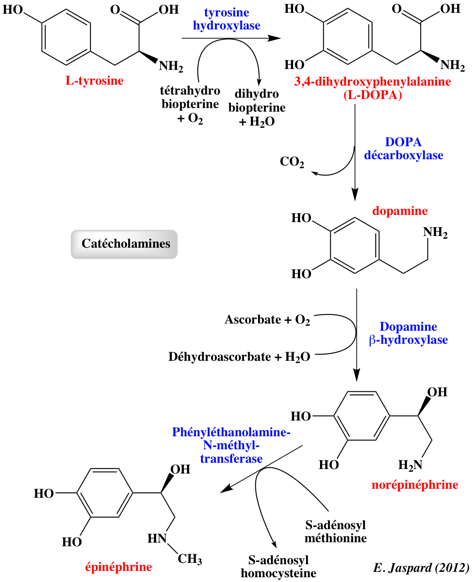
6. Action de l'adrénaline (appelée aussi épinéphrine) L'adrénaline (ou épinéphrine) est une hormone et un neurotransmetteur.
C'est une catécholamine dérivée de la tyrosine [(dihydroxy-3,4 phényl)-1 méthylamino-2 éthanol]. La dernière étape met en jeu la S-adénosylméthionine.
L'adrénaline est sécrétée par les glandes surrénales en réponse à des signaux nerveux. L'un de ces effets est d'accélèrer la dégradation du glycogène. Il y a alors une augmentation du taux intracellulaire de glucose 6-phosphate ce qui amène la glycolyse à s'accélérer dans le muscle et induit le foie à libérer plus de glucose dans le sang. Beaucoup de tissus sont sensibles à l'adrénaline. Le récepteur β-adrénergique de l'adrénaline stimule la protéine G appelée GS qui stimule à son tour l'activité de l'adénylate cyclase (formation d'AMPc). L'AMPc accélère la dégradation du glycogène. Cet effet est identique à celui observé par la fixation du glucagon à son récepteur. En revanche, la fixation de l'adrénaline au récepteur α1-adrénergique met en jeu un système de régulation différent qui fait intervenir la phospholipase C. Le diacylglycérol active la protéine kinase C. L'une des cibles de cette protéine kinase C est le récepteur de l'insuline. Celui-ci a peu d'affinité à l'état phosphorylé pour l'insuline.
L'action de l'adrénaline est donc d'atténuer l'effet de l'insuline. L'insuline qui, par ailleurs active la transcription du gène codant la glucokinase. |
| 7. Liens Internet et références bibliographiques |
| Nadeau et al. (2005) "Cryoelectron microscopy reveals new features in the three-dimensional structure of phosphorylase kinase" Protein Science 14, 914 - 920 | Article |
|
"Glycogen breakdown (glycogenolysis) [Homo sapiens]" (Base de données : "Reactome") |
Aller au site |
![]()