| Modèle allostérique de Jacques MONOD, Jeffries WYMAN & Jean-Pierre CHANGEUX (1965) |
| Tweet |
|
|
1. Introduction au modèle MWC ou modèle concerté ou de "tout ou rien" 2. Les bases du modèle MWC 3. Nomenclature du modèle MWC |
|
1. Introduction au modèle MWC ou modèle concerté ou de "tout ou rien" (voir une partie de l'article original) a. L'un des moyens employés par la cellule pour contrôler le flux global d'une voie métabolique est la modulation de l'activité de certaines enzymes selon un mécanisme extrêmement sophistiqué : la régulation allostérique :
Voici deux extraits de l'introduction de l'article de Jacques Monod, Jeffries Wyman & Jean-Pierre Changeux (1965) :
b. Le terme "allostérique" vient des deux mots Grecs : "allos" et "stereos" qui signifient : "une autre forme" ou "un autre solide". Il peut donc s'entendre comme "une autre conformation". Il fût employé par J. Monod & François Jacob (1961) pour désigner le processus d'inhibition du précurseur du Trp par le Trp lui-même Avec le développement de modèles décrivant les effets allostériques, il a pris un sens supplémentaire : "un autre site". c. Les effecteurs allostériques sont des molécules autres que les substrats (en général) et dont la fixation en des sites distincts (voire identiques, compétitifs) des sites de fixation des substrats diminue l'activité de l'enzyme (inhibiteurs) ou l'augmente (activateurs). Les effecteurs sont souvent :
|
2. Les bases du modèle MWC a. Deux types d'effets allostériques :
b. Les propriétés générales des systèmes allostériques sont les suivantes :
c. Les hypothèses du modèle MWC
|
La partie qui suit est inspiré du livre "Enzyme Kinetics" (1993) de Irwin H. SEGEL (Wiley & sons. ed). |
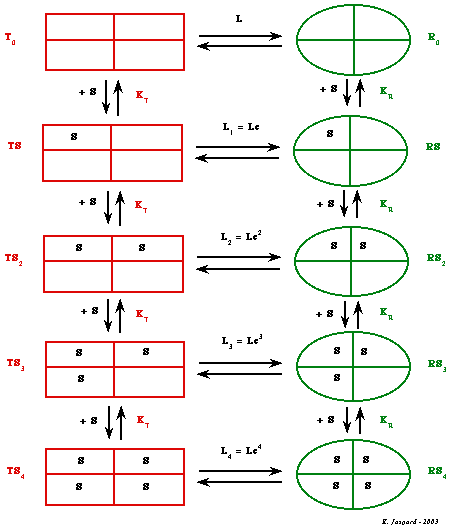
3. Nomenclature du modèle MWC L'enzyme étudiée par Monod, Wyman & Changeux pour établir leur modèle est la phosphofructokinase, enzyme tétramérique constituée de 4 sous-unités identiques. Les deux états conformationnels que peut adopter un protomère sont dénommés respectivement :
Ces deux conformations sont en équilibre (quelle que soit la concentration d'un ligand donné) et cet équilibre est régit par la constante allostérique L0 : L0 |
Schéma général de la fixation successive de 4 molécules de substrat "...by reason of symmetry and because the binding of any one ligand molecule is assumed to be intrinsically independent of the binding of any other, the microscopic dissociation constants are the same ..." Le modèle MWC est symétrique et il s'appuie sur l'hypothèse que la fixation d'une quelconque molécule de substrat (ou ligand) est indépendante de la fixation d'une autre molécule de substrat (ou ligand).
En conséquence, les constantes de dissociation microscopiques sont les mêmes pour tous les sites de fixation homologues dans chacun des 2 états (R & T). On adopte donc la nomenclature suivante : KT = KT0 = KTS = KTS2 = KTS3 = KTS4 KR = KR0 = KRS = KRS2 = KRS3 = KRS4 |
L'entropie joue un rôle dans la coopérativité :
L'entropie S est donc plus élevée pour la fixation de la première molécule de substrat : S = k . log W
La première molécule de substrat doit donc surmonter une plus grande variation d'entropie que la dernière molécule de substrat : l'affinité des sites de fixation est apparemment augmentée après la fixation d'une molécule de ligand à un site de fixation. C'est la coopérativité positive qui se traduit par une sigmoïde. |
![]()