| L'épissage des ARN messager pré-matures : le spliceosome |
| Tweet |
|
|
1. Introduction 2. Structures des complexes ribonucléoprotéiques snRNP 3. Composition des complexes ribonucléoprotéiques snRNP et auxilliaires (non snRNP) 4. Mécanismes au sein du spliceosome
|
5. Un mécanisme évolutif commun au spliceosome et aux introns du groupe II (ribozyme) ? 6. Le spliceosome U12-dépendant 7. Les protéines SR ("Serine-arginine-Rich splicing factors") 8. La protéine Prp8 9. Liens Internet et références bibliographiques |
| 1. Introduction
Chez les Eucaryotes, les génes sont transcrits sous forme d'ARN messagers pré-matures (synonymes : transcrits primaires ou pré-ARNm) qui contiennent :
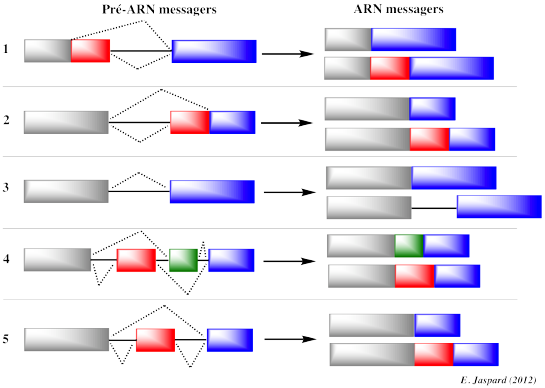
Les introns sont non retenus : ils sont clivés selon un mécanisme appelé épissage qui peut être constitutif (le même intron est systématiquement clivé) ou alternatif. Remarque : Les introns du groupe II sont des introns auto-catalytiques ou ribozymes. Les exons sont assemblés selon différentes combinaisons forment la séquence finale qui code la protéine ou l'ARN. Richard Roberts et Phillip Sharp ont obtenu le Prix Nobel en 1993 pour la découverte de la structure des gènes [introns - exons] et le principe de l'épissage. Figure ci-dessous : 5 types d'épissage alternatif
Ne sont pas représentés les épissages :
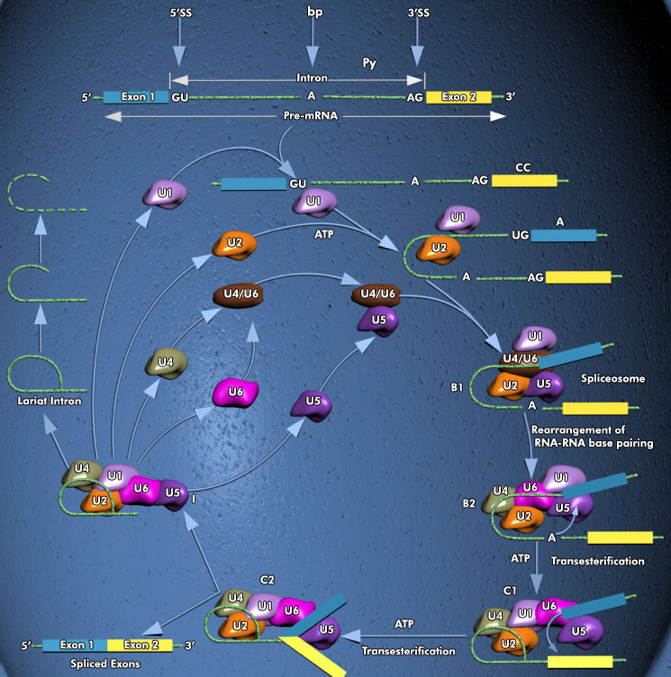
Les introns sont présents dans le génome de tous les Eucaryotes séquencés à ce jour . De multiples introns interrompent les séquences codantes de la très grande majorité des gènes des animaux et des plantes. En revanche, la densité des introns chez les fungi et les Eucaryotes unicellulaires est hautement variable : beaucoup n'en contiennent que quelques exemplaires dans l'ensemble de leur génome. Chez les Eucaryotes, le phénomène d'épissage (clivage - coupure : "splicing") des introns est effectué par un supra-complexe ribonucléoprotéique appelé le spliceosome : multi-complexes d'ARN et de protéines de plusieurs millions de Da. Cette maturation des ARNm (qui inclue également l'addition de la coiffe en 5', la poly-adénylation en 3' et l'édition) au sein des spliceosomes transforme les pré-ARNm en ARNm matures fonctionnels. Les ARNm matures sont ensuite transportés du noyau vers le cytoplasme via les pores nucléaires pour y être traduits. Figure ci-dessous : Ensemble des mécanismes de l'épissage d'un pré-ARNm ("Pre-mRNA").
Source figure : Qiagen Il existe 2 types de spliceosomes chez la plupart des Eucaryotes :
Le décryptage complet et précis des mécanismes de l'épissage et de leur régulation est d'une importance capitale pour des approches thérapeutiques. En effet, un grand nombre de pathologies et de maladies génétiques chez l'homme sont liées à des phénomènes d'épissage aberrants des pré-ARNm. |
|
2. Structures des complexes ribonucléoprotéiques snRNP Beaucoup de protéines du spliceosome ont été cristallisées. On dipsose donc d'un ensemble quasi-complet d'informations pour le décryptage à l'échelle moléculaire des mécansimes structuraux, d'association et de catalyse du spliceosome et de l'épissage des introns. L'épissage par le spliceosome est une suite d'associations [ARN-ARN], [protéines-protéines] et [ARN-protéines] extrêmement dynamiques. Plusieurs méga-complexes ribonucléoprotéiques se forment et se dissocient au cours du temps. Ce sont les complexes E, A, pre-B ("pre-B complex"), B ("pre-catalytic spliceosome"), Bact ("activated spliceosome"), B* ("catalytically activated spliceosome"), C ("catalytic step I complex"), C* ("step II catalytically activated complex"), P ("post-catalytic spliceosome") et ILS ("Intron Lariat Spliceosome").
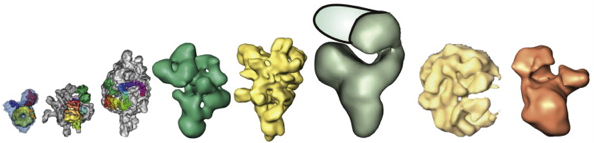
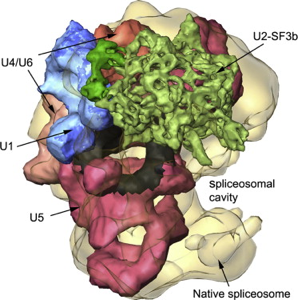
Source : KEGG En ce qui concerne les complexes assemblés, ils sont si gros que seule la cryo-microscopie électronique (cryo-EM) est apte, actuellement, à en déterminer la structure (figure ci-dessous).
Source : Lührmann & Stark (2009) De gauche à droite : U1 snRNP - SF3b - U11/U12 di-snRNP - U5 snRNP - U4/U6.U5 tri-snRNP - spliceosome BDU1 - spliceosome natif - complexe C |
| Résumé des structures déterminées par diverses techniques | ||||
| Complexe | Méthode | résolution | Année | Référence |
| snRNP U1 | Cryo-EM | 10 Å | 2001 | Stark et al. (2001) "Arrangement of RNA and proteins in the spliceosomal U1 small nuclear ribonucleoprotein particle" Nature 409, 539 - 542 |
| "Splicing factor 3b" (SF3b) - groupe de 7 protéines de U2 snRNP | Cryo-EM | 10 Å | 2003 | Golas et al. (2003) "Molecular architecture of the multiprotein splicing factor SF3b" Science 300, 980 - 984 |
| complexe BΔU1 | Cryo-EM | 40 Å | 2004 | Boehringer et al. (2004) "Three-dimensional structure of a pre-catalytic human spliceosomal complex B" Nat. Struct. Mol. Biol. 11, 463 - 468 |
| di-snRNP U11/U12 | Cryo-EM | 12 Å | 2005 | Golas et al. (2005) "Major conformational change in the complex SF3b upon integration into the spliceosomal U11/U12 di-snRNP as revealed by electron cryomicroscopy" Mol. Cell 17, 869 - 883 |
| snRNP U5 | Cryo-EM | 26 - 32 Å | 2006 | Sander et al. (2006) "Organization of core spliceosomal components U5 snRNA loop I and U4/U6 Di-snRNP within U4/U6.U5 Tri-snRNP as revealed by electron cryomicroscopy" Mol. Cell 24, 267 - 278 |
| di-snRNP U4/U6 | 40 Å | |||
| tri-snRNP U4/U6.U5 | 19 - 24 Å | |||
| complexe A | Cryo-EM | 40 - 50 Å | 2007 | Behzadnia et al. (2007) "Composition and three-dimensional EM structure of double affinity-purified, human prespliceosomal A complexes" EMBO J. 26, 1737 - 1748 |
| complexe C | Cryo-EM | 29 Å | 2007 | Ohi et al. (2007) "Structural characterization of the fission yeast U5.U2/U6 spliceosome complex" Proc. Natl. Acad. Sci. USA 104, 3195 - 3200 |
| coeur catalytique de U1 snRNP | Cristallographie | 5,5 Å | 2009 | Pomeranz Krummel et al. (2009) "Crystal structure of human spliceosomal U1 snRNP at 5.5 A resolution" Nature 458, 475 - 480 |
| snRNP U4 (anneau heptamèrique de protéines Sm + AUUUUUG) | Cristallographie | 3,6 Å | 2011 | Leung et al. (2011) "Structure of the spliceosomal U4 snRNP core domain and its implication for snRNP biogenesis" Nature 473, 536 - 539 |
spliceosome
|
Modélisation In silico | ----- | 2012 | Frankenstein et al. (2012) "A Unique Spatial Arrangement of the snRNPs within the Native Spliceosome Emerges from In Silico Studies" Structure 20, 1097 - 1106 |
tri-snRNP U4/U6.U5 - Saccharomyces cerevisiae
|
Cryo-EM | 5,9 Å | 2015 | Nguyen et al. (2015) "The architecture of the spliceosomal U4/U6.U5 tri-snRNP" Nature 523, 47 - 52 |
| 3,7 Å | 2016 | Nguyen et al. (2016) "Cryo-EM structure of the yeast U4/U6.U5 tri-snRNP at 3.7 Å resolution" Nature 298 - 302 | ||
complexe C* Homo sapiens : complexe C activé après action de l'ARN hélicase PRP16 et avant l'étape 2 catalyse |
Cryo-EM | 5,9 Å | 2017 | Bertram et al. (2017) "Cryo-EM structure of a human spliceosome activated for step 2 of splicing" Nature 542, 318 - 323 |
complexe C* Saccharomyces cerevisiae : complexe C activé après l'étape 2 catalyse |
Cryo-EM | 4,0 Å | 2017 | Yan et al. (2017) "Structure of a yeast step II catalytically activated spliceosome" Science 355, 149 - 155 |
| 17S U2 snRNP (homme) - principale sous-unité des complexes E et A | Cryo-EM | PDB 6Y5Q 7,1 Å |
2020 | |
Bien que beaucoup de protéines du spliceosome aient été cristallisées, près de la moitié des séquences des protéines constitutives du spliceosome de l'homme sont prédites intrinsèquement désordonnées ("intrinsically disordered" - voir Korneta & Bujnicki, 2012). La distribution [ordre/désordre] au sein des protéines du spliceosome est irrégulière. En particulier, les protéines impliquées dans la reconnaissance des structures secondaires (reconnaissance de pré-ARNm, définition d'intron et définition d'exon, assemblage et dynamique du spliceosome - voir ci-dessous), sont plus désordonnées que les protéines impliquées dans l'assistance à la catalyse de l'épissage. Les régions désordonnées conservées des protéines du spliceosome sont plus récentes du point de vue de l'évolution et moins étendues que les régions ordonnées, ce qui suggère que les régions désordonnées ont été ajoutées à un "coeur" ordonnée pré-existant. Le protéome du spliceosome est prédit contenir un taux global de désordre plus élevé que celui des ribosomes (autres particules ribonucléoprotéiques massives), ce qui est cohérent avec les fonctions distinctes des protéines constitutives de ces 2 types de complexes. |
3. Composition des complexes ribonucléoprotéiques snRNP et auxilliaires (non snRNP) Nomenclature :
Le spliceosome est caractérisé par de grandes modifications de sa composition en protéines (échange très dynamique) et de son réseaux d'interactions entre les snRNP et le pré-ARNm. Chaque snRNP est composé :
La machinerie protéique des spliceosomes est donc colossale et dynamique (voir tableau ci-dessous) . Chez l'homme, on dénombre :
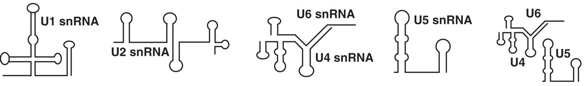
Les protéines des spliceosomes sont très phosphorylées et subissent d'autres types de modifications post-traductionnelles. Les spliceosomes de la levure contiennent un nombre plus réduit de protéines (mais cependant tout à fait conséquent). Figure ci-dessous : Structure secondaire des snRNA des complexes snRNP du spliceosome majeur de l'homme.
Source : Will & Lührmann (2011) |
| Composition en protéines des complexes snRNP et auxilliaires du spliceosome majeur de l'homme | ||||||
| U1 (12 Svedberg) | U2 (17 S) | U4/U6 (13 S) | U5 (20 S) | Prp19 | EJC/TREX | Composants communs |
|
|
|
|
|
|
|
U1 related proteins
|
U2 related proteins
|
U4/U6.U5 tri-snRNP associated proteins
|
Prp19 related proteins
|
|||
Les protéines effectrices Prp2, Prp5, Prp16, Prp17, Prp18, Prp22, Prp43, Slu7 et UAP56 interviennent également au cours de l'épissage par le spliceosome. |
||||||
| Aller à KEGG : tous les spliceosomes (menu déroulant du haut) avec figures interactives qui renvoient vers toutes les informations concernant chaque RNA et protéines. | ||||||
|
Détails pour U1 (les liens renvoient vers les informations de KEGG) :
|
Détails pour U2 (les liens renvoient vers les informations de KEGG ou Uniprot) :
|
La spectromètrie de masse a permis d'étudier en détail :
La base de données "Spliceosome Database" :
Modifications post-traductionnelles Au moins 4 protéines kinases phosphorylent un très grand nombre de protéines du spliceosome : SR protéine kinase 1, SR protéine kinase 2, Prp4 kinase et Clk/Sty kinase.
|
|
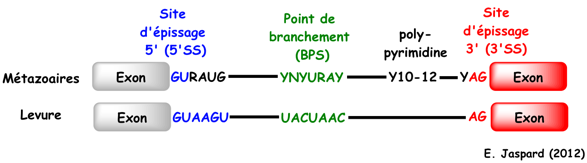
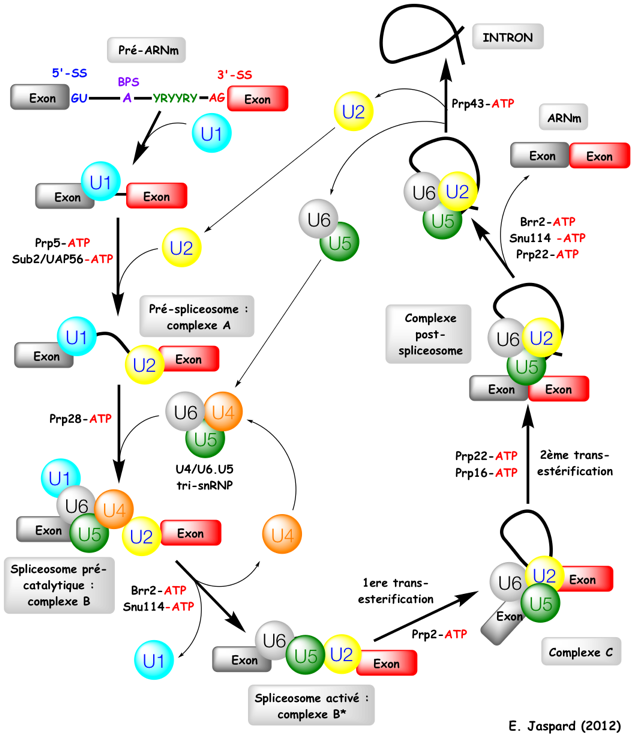
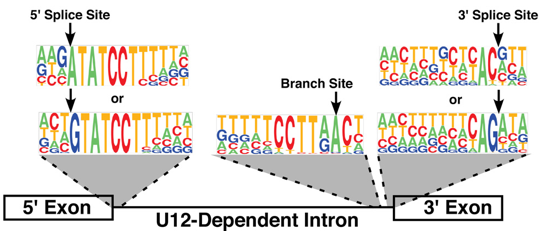
4. Mécanismes au sein du spliceosome La majorité des spliceosomes reconnaissent les introns qui commencent par 5'-GU et se terminent par AG-3'. Ils reconnaissent aussi plusieurs séquences consensus clé (éléments agissant en cis-). Certains spliceosomes reconnaissent les introns qui commencent par 5'-AU et se terminent par AC-3'. Ils utilisent les snRNP U11, U12, U4atac-U6atac. Figure ci-contre : Séquences consensus de pré-ARNm subissant un épissage. BPS : "branch point sequence" - 5'SS : "5' splicing site" - 3'SS : "3' splicing site" - N : n'importe quel nucléotide - R : une purine - Y : une pyrimidine
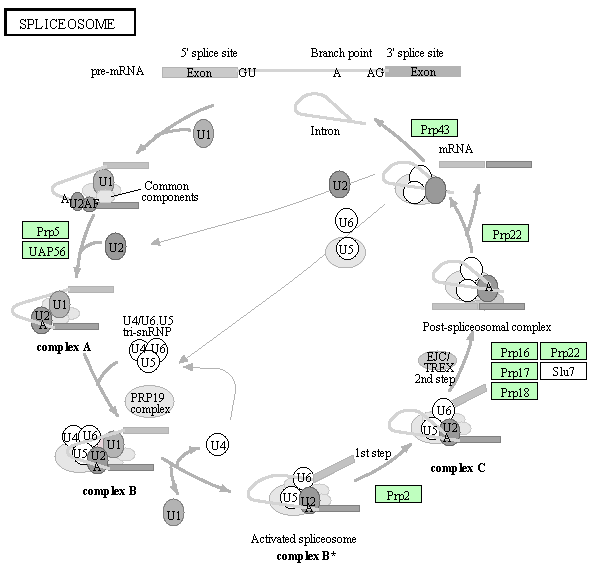
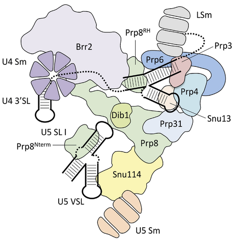
a. Etapes de l'épissage et formation des complexes successifs Seul un petit nombre de protéines effectrices qui interviennent dans le processus complet de l'épissage sont indiquées dans la figure ci-dessous.
Remarque : Prp5, Prp16 et Prp22 ont également une fonction de correction ("proofreading").
Etape 1 : Le snRNP U1 reconnait le site 5'SS du pré-ARNm. Les protéines SF1 et l'héterodimère U2AF ("U2 Auxilliary Factor") se fixent au BPS et à la séquence poly-pyrimidine, respectivement (non montré dans la figure ci-contre). SF1/BBP interagit avec U2AF65 via son domaine RRM ("RNA Recognition Motif" - voir la "définition d'exon") et U2AF65 se fixe sur le dinucléotide AG du site 3'SS. Ces interactions aboutissent à la formation du complexe E. Etape 2 : Le snRNP U2 se fixe sur le BPS du pré-ARNm, formant ainsi le complexe A. Etape 3 : Le snRNA U4 et le snRNA U6 établissent un grand nombre de liaisons.
Le complexe A s'associe au tri-snRNP préformé U4/U6.U5 pour aboutir au complexe pré-catalytique B : il contient l'ensemble complet [U1, U2, U4/U6 et U5 et les 5 snRNA]. La protéine Prp28 (ATPase/hélicase ARN-dépendante de type DExD/H) transfère le site 5′SS de l'extrémité 5′ du snRNA de U1 vers une séquence conservée ACAGAGA du snRNA de U6. Cet évènement initie l'activation pour la catalyse du complexe B. Etape 4 : Le duplex U4/U6 est déroulé par Brr2. Le complexe B est alors activé pour la catalyse par :
Brr2 (EC 3.6.4.13) :
Régulations croisées :
Ce remodelage aboutit au spliceosome activé : le complexe B*. Etape 5 : Le complexe B* catalyse la 1ère réaction de trans-esterification. Etape 6 : On aboutit au complexe C qui catalyse la 2ème réaction de trans-esterification. Etape 7 : L'ARNm (exons associés) est relargué du complexe post-spliceosome sous forme d'une particule ribonucléoprotéique (protéines associées). Etape 8 : L'intron destiné à la dégradation est relargué par dissociation de U2, U5 et U6. Prp43 fonctionne pendant le désassemblage du spliceosome. Elle forme le complexe NTR avec les protéines Ntr1/Spp382 et Ntr2. |
|
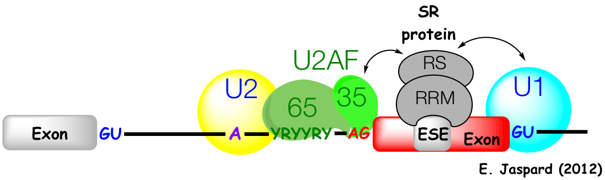
Quand un intron fait plus de 200 - 250 nucléotides (ce qui est le cas de nombreux introns chez les Eucaryotes supérieurs), les complexes d'épissage se forment d'abord autour d'un exon. Le snRNP U1 se fixe au site 5'SS de l'exon situé en 5' (figure ci-dessous). Cette fixation induit l'association des 2 sous-unités du facteur auxilliaire de U2 ("U2 Auxilliary Factor" - U2AF65 / 65 KDa et U2AF35 / 35 KDa) avec la séquence poly-pyrimidine ("polypyrimidine tract recognition") en amont du site 3'SS de ce même exon. La sous-unité U2AF35 établit des liaisons spécifiques avec le site 3'SS. Figure ci-dessous : Complexes d'épissage formés le long des introns lors des premières étapes de l'assemblage du spliceosome.
Le snRNP U2 est à son tour recruté et se fixe sur BPS en amont de cet exon. Une protéine SR ("SR protein") contenant un domaine riche en dipeptide [Arg / Ser] (RS) et un motif de reconnaissance de l'ARN ("RNA Recognition Motif" - RRM) interagit avec des séquences d'activation de l'épissage ("Exonic Splicing Enhancer" - ESE) au sein de cet exon Les protéines SR établissent un réseau d'interactions protéine - protéine autour de l'exon. Ces interactions stabilisent l'ensemble [complexes snRNP - exon]. |
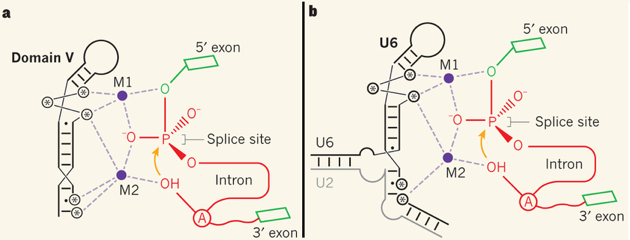
5. Un mécanisme évolutif commun au spliceosome et aux introns du groupe II (ribozyme) ? Le splicéosome possède des ions magnésium dans son site actif qui sont des cofacteurs catalytiques. On cherche depuis longtemps à savoir si les ligands de ces ions métalliques sont fournis par les snRNA (ARN spécifique de chaque snRNP) ou par les protéines des complexes ribonucléoprotéiques snRNP, car la réponse à cette question a des incidences sur l'aspect évolutif du mécanisme d'épissage. Figure ci-dessous - partie a : les introns du groupe II sont des introns auto-catalytiques ou ribozymes.
Source : Strobel (2013)
Les ARN des introns du groupe II contiennent une structure en épingle à cheveux appelée domaine V. Les réactions de transfert de phosphoryle lors de l'épissage (dont la première est indiquée par une flèche orange) sont catalysées par deux ions métalliques divalents (M1 et M2) qui établissent des liaisons de coordination (pointillés violets) avec :
Figure ci-dessus - partie b : analogie avec le snRNA U6 Un travail récent (Fica et al., 2013) a montré que le snRNA de U6 (qui forme une structure en épingle à cheveux semblable à celle du domaine V) du spliceosome de la levure fournit un très grand nombre d'atomes d'oxygène de groupements phosphate spécifiques qui se lient aux deux ions métalliques catalytiques du site actif, de manière similaire à la coordination des ions métalliques des introns de groupe II. Ces résultats :
Il y a plus de 100 protéines dans le splicéosome: quels rôles ont ces protéines si l'ARN est le catalyseur de l'épissage de l'ARN pré-messager ?
|
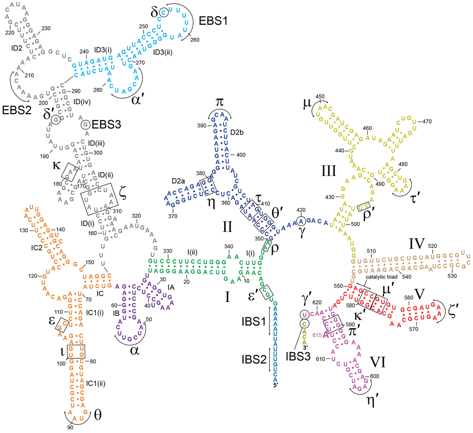
Rôle du lariat dans l'épissage des introns de groupe II Des élements de réponse aux questions qui précèdent sont apportés progressivement. Par exemple, en 2014, Robart et al. (2014) ont obtenu la première structure (à une résolution 3.7 Å) d'un intron du groupe II sous sa forme lariat post-catalytique avec l'exon (produit) lié. C'est la première structure d'une molécule d'ARN 2′-5′. Figure ci-dessous : structure de l'intron du groupe IIB P.li.LSUI2 de l'algue brune Pylaiella littoralis.
Source : Robart et al. (2014) Il existe 3 classes structurales d'introns du groupe II (IIA, IIB et IIC). Les introns du groupe II possèdent la séquence consensus GUGYG (avec Y = U ou C). Les interactions tertiaires sont indiquées par des lettres grecques et les domaines par des chiffres romains.
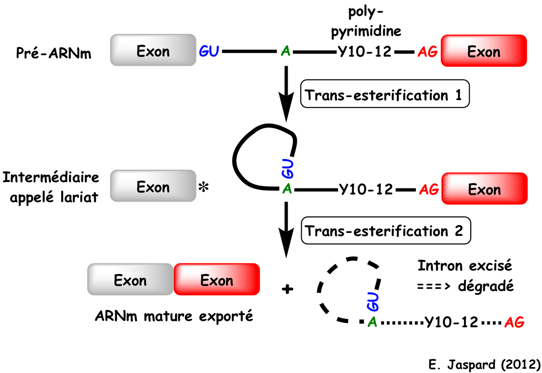
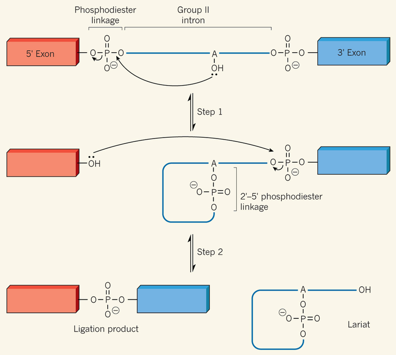
Les enzymes ARN ou ribozyme appelés introns de groupe II catalysent leur propre retrait de l'ARN et joignent les séquences exoniques flanquantes.
Source : R. Batey (2014) Lors de la première étape de l'épissage, le groupe hydroxyle d'une adénosine (A) du BPS de l'intron attaque la liaison phosphodiester du site 5'SS, ce qui forme :
Lors de la deuxième étape, le groupe hydroxyle 3′ de l'exon 5′ attaque la liaison phosphodiester sur le site 3'SS, ce qui forme :
|
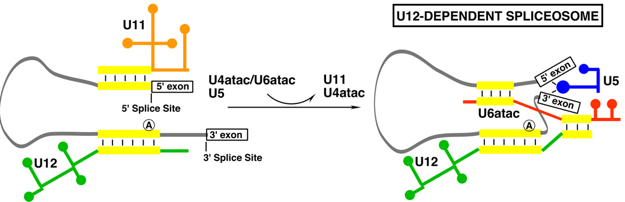
6. Le spliceosome U12-dépendant Quelques espèces de métazoaires et de plantes possède un second spliceosome dit "mineur" : le spliceosome U12-dépendant qui catalyse l'épissage des introns rares de type U12 ("U12-type introns"). Le spliceosome U12-dépendant est constitué de snRNP dont la composition est différente, mais dont le fonctionnement est semblable, au spliceosome "majeur" U2-dépendant. Il contient les snRNP U11, U12 et U4atac/U6atac. Le snRNP U5 est commun aux 2 types de spliceosome. Voir quelques snRNA U11, U12, U4atac et U6atac. Les introns de type U12 ont des séquences consensus différentes des introns de type U2 :
Source : Padgett (2012)
Au cours de la formation du spliceosome U12-dépendant, les snRNPs U11 et U12 forment un complexe di-snRNP U11/U12. Sept protéines (20K, 25K, 31K, 35K, 48K, 59K et 65K) s'associent à ce di-snRNP. [U11/U12-65K], [U11/U12-59K] et [U11/U12-48K] sont nécessaires pour la sélection du site 5′SS et du BPS.
Source : Padgett (2012) |
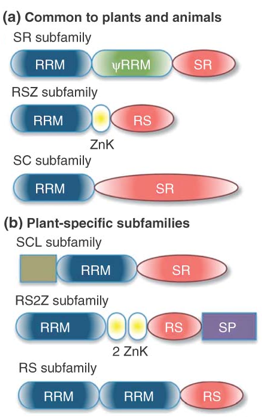
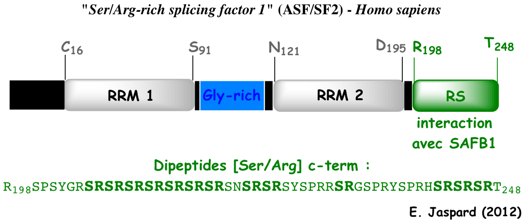
7. Les protéines SR ("Serine-arginine-Rich splicing factors") La détection / sélection des sites d'épissage est régulée par la fixation initiale de facteurs d'épissage riche en Ser/Arg ("Serine-arginine-Rich splicing factors" - protéines SR) au pré-ARNm. Les protéines SR ont généralement une localisation nuclèaire. Cependant, certaines d'entre elles (exemples : ASF/SF2, 9 G8, and SRp20) sont aussi des transporteurs d'ARNm entre le noyau et le cytoplasme. L'activité des protéines SR est finement régulée par phosphorylation / déphosphorylation. Les protéines SR contiennent :
Source : Barta et al. (2010)
Voir toutes les protéines SR ("splicing factor SR family" - Uniprot). Les activateurs et suppresseurs de l'épissage ("exon-splicing enhancers / silencers") sont reconnus par des sous-ensembles spécifiques de protéines SR : SRp20, SRp30a (ASF/SF2) SRp30c, 9 G8, SRp40, SRp55, SRp70, SRp75, et SC35. La teneur respective de différentes protéines SR et la présence d'activateurs / suppresseurs de l'épissage influencent l'assemblage ultérieur des protéines du spliceosome. |
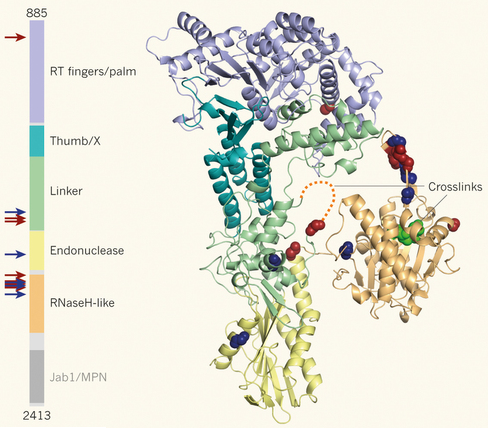
8. La protéine Prp8 ("Pre-mRNA Processing factor 8") La protéine Prp8 est la plus grosse protéine (274 KDa chez l'homme) du spliceosome (2335 et 2413 acides aminés chez l'homme et la levure, respectivement) et la plus conservée (62%d' identité entre l'homme et la levure). Chez l'homme, Prp8 est un élément des 2 types de spliceosome (U2- et U12-dépendants) : elle fait partie du snRNP U5, du tri-snRNP U5.U4/U6 et du tri-snRNP U5.U4atac/U6atac et du complexe EJC ("mRNA splicing-dependent Exon Junction Complex"). Prp8 est considérée comme l'un des régulateurs clé du fonctionnement du spliceosome. En effet, elle régule l'activité de protéines clé de l'épissage (formation du complexe B activé) : notamment celle de l'hélicase Brr2 et de la GTPase Snu114 (qui régule elle-même l'activité de Brr2). Prp8 interagit avec :
Prp8 possède :
Source : Query & Konarska (2013)
La structure de Prp8 (résidus 885–2413) de la levure (complexée à Aar2 - facteur d'assemblage du snRNA U5) révèle des domaines étroitement associés qui ressemblent à la transcriptase inverse (bactérienne) d'introns du groupe II et à une endonucléase de restriction de type II (Galej et al., 2013). La structure permet de mieux comprendre l'architecture du site actif du spliceosome et renforce l'hypothèse que l'épissage des ARN pré-messagers nucléaires et l'épissage des des introns de groupe II ont une origine évolutive commune. |
| 9. Liens Internet et références bibliographiques |
|
Qiagen : "RNAi pathway" Animation très didactique du fonctionnement du spliceosome "RNA Splicing" |
|
|
KEGG : tous les spliceosomes (menu déroulant du haut) avec figures interactives qui renvoient vers toutes les informations concernant chaque RNA et protéines. "Spliceosome Database : A source of information for the Spliceosome" "U12DB : a database of orthologous U12-type spliceosomal introns" "Rfam : The Rfam database is a collection of RNA families" "SpliProt3D : an online database of structures of proteins of the human spliceosome" |
|
|
Luhrmann & Stark (2009) "Structural mapping of spliceosomes by electron microscopy" Curr. Opin. Struc. Biol. 19, 96 - 102 Korneta & Bujnicki (2012) "Intrinsic Disorder in the Human Spliceosomal Proteome" PLoS Comput. Biol. 8, e1002641 Korneta et al. (2012) "Structural bioinformatics of the human spliceosomal proteome" Nucleic Acids Res. 40, 7046 - 7065 Bolisetty & Beemon (2012) "Splicing of internal large exons is defined by novel cis-acting sequence elements" Nuc. Acids. Res. 40, 9244 - 9254 |
|
Fabrizio et al. (2009) "The Evolutionarily Conserved Core Design of the Catalytic Activation Step of the Yeast Spliceosome" Mol. Cell 36, 593 - 608 Will & Lührmann (2011) "Spliceosome Structure and Function" Cold Spring Harb. Perspect. Biol. 3 Frankenstein et al. (2012) "A Unique Spatial Arrangement of the snRNPs within the Native Spliceosome Emerges from In Silico Studies" Structure 20, 1097 - 1106 Schmitzova & Pena (2012) "Emerging views about the molecular structure of the spliceosomal catalytic center" RNA Biol. 9 Burke et al. (2012 ) "Structure of the yeast U2/U6 snRNA complex" RNA 18, 673 - 683 |
|
|
Rylond B. (2007) "Targeting the spliceosome" Nat. Chem. Biol. 3, 533 - 535 Wahl et al. (2009) "The Spliceosome: Design Principles of a Dynamic RNP Machine" Cell 136, 701 - 718 Barta et al. (2010) "Implementing a Rational and Consistent Nomenclature for Serine/Arginine-Rich Protein Splicing Factors (SR Proteins) in Plants" Plant Cell 22, 2926 - 2929 Wachter et al. (2012) "The Role of Polypyrimidine Tract-Binding Proteins and Other hnRNP Proteins in Plant Splicing Regulation" Front. Plant Sci. 3, 81 Padgett (2012) "New connections between splicing and human disease" Trends Genet. 28, 147 - 154 |
|
|
U12 minor spliceosomal RNA : RFAM Family: U12 (RF00007) Allioto, TS. (2007) "U12DB: a database of orthologous U12-type spliceosomal introns" Nucleic Acids Res. D110 - D115 Davila Lopez et al. (2008) "Computational screen for spliceosomal RNA genes aids in defining the phylogenetic distribution of major and minor spliceosomal components" Nucleic Acids Res. 36, 3001 - 3010 Strobel (2013) "Biochemistry: Metal ghosts in the splicing machine" Nature 503, 201-202 Fica et al. (2013) "RNA catalyses nuclear pre-mRNA splicing" Nature 503, 229-234 |
|
|
Query & Konarska (2013) "Structural biology: Spliceosome's core exposed" Nature 493, 615 - 616 Galej et al. (2013) "Crystal structure of Prp8 reveals active site cavity of the spliceosome" Nature 493, 638 - 643 R. Batey (2014) "Structural biology: Lariat lessons" Nature 514, 173-174 Robart et al. (2014) "Crystal structure of a eukaryotic group II intron lariat" Nature 514, 193-197 |
|
|
Nguyen et al. (2015) "The architecture of the spliceosomal U4/U6.U5 tri-snRNP" Nature 523, 47 - 52 Nguyen et al. (2016) "Cryo-EM structure of the yeast U4/U6.U5 tri-snRNP at 3.7 Å resolution" Nature Yan et al. (2017) "Structure of a yeast step II catalytically activated spliceosome" Science 355, 149 - 155 Bertram et al. (2017) "Cryo-EM structure of a human spliceosome activated for step 2 of splicing" Nature 542, 318 - 323 |
|
|
Zhang et al. (2019) "Structures of the human spliceosomes before and after release of the ligated exon" Cell Res. 29, 274 - 285 Li et al. (2019) "A unified mechanism for intron and exon definition and back-splicing" Nature 573, 375 - 380 Zhang et al. (2020) "Molecular architecture of the human 17S U2 snRNP" Nature 583, 310 - 313 |
![]()