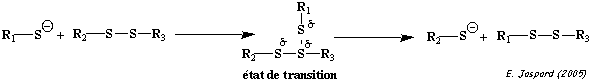| Insuline : cystéine et pont disulfure |
| Tweet |
|
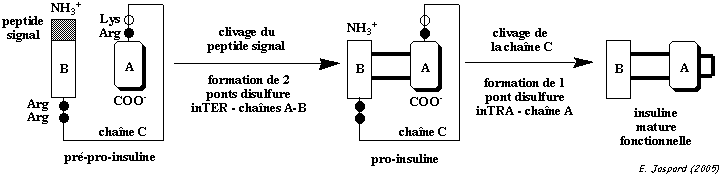
L'insuline est synthétisée sous la forme d'un précurseur, la pré-pro-insuline dans les cellules β des îlots de Langerhans.
Comment ces données sont-elles en accord avec l'idée que la séquence primaire d'une protéine dicte son repliement ? |
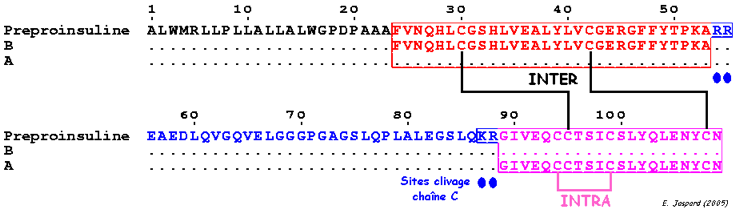
1. Maturation de l'insuline Deux ponts disulfure INTER-chaînes de l'insuline se forment dès la biosynthèse de la pro-insuline. La chaîne C de la pro-insuline a pour rôle de positionner correctement les chaînes A et B. Quand la pro-insuline est correctement repliée, le peptide C est éliminé et un pont disulfure INTRA-chaîne se forme. L'insuline adopte alors sa forme fonctionnelle.
Cette maturation par l'élimination du peptide C retarde l'apparition de l'activité hormonale de l'insuline jusqu'à ce qu'elle soit empaquetée dans les granules de sécrétion.
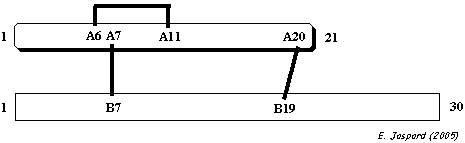
Pont INTRA-chaîne : CYS A 6 - CYS A 11 - Ponts INTER-chaînes : CYS A 7- CYS B 7 / CYS A 20- CYS B 19
|
|
Visualisation de l'insuline de porc à une résolution de 1,5 Å Code PDB : 4INS Les 4 chaînes (A, B, C et D) sont colorées.
|
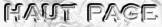
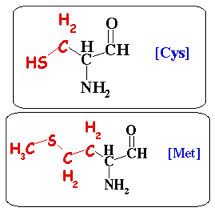
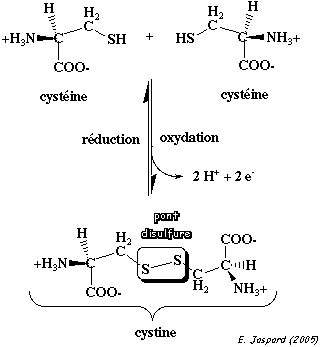
2. La cystéine et la méthionine La cystéine (C ou Cys ou acide L-2-amino-3-mercaptopropionique) est codée par les 2 codons UGU et UGC. Sa chaîne latérale porte un groupement sulfhydryle (-SH) extrêmement réactif dont le pKa = 8.33.
L'autre acide aminé sulfuré, la méthionine (M ou Met) , porte un groupe méthyle supplémentaire. La méthionine est plus hydrophobe, d'un encombrement stérique plus important et moins réactive.
|
|
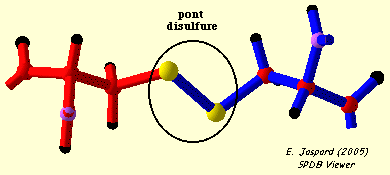
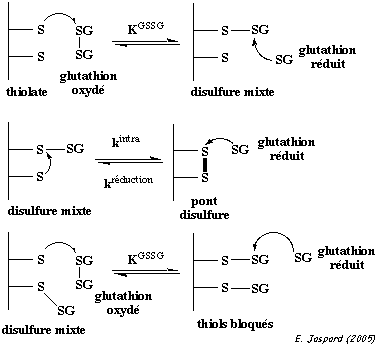
3. Formation d'un pont disulfure a. Introduction Un pont disulfure est une liaison covalente entre 2 atomes de soufre de la chaîne latérale de 2 résidus cystéines (Cys) réduits. Les ponts disulfures sont formés dans un environnement cellulaire oxydant. Le cytoplasme n'étant pas un milieu oxydant, il y a trés peu de protéines intracellulaires qui possèdent des ponts disulfures. Toutes les cystéines d'une protéine ne forment pas un pont disulfure :
Un pont disulfure peut être établi entre 2 cystéines :
Le nombre de ponts disulfures (intra et/ou inter) au sein des protéines varie trés grandement : les toxines, bien qu'étant de petites protéines (20 à 30 acides aminés) peuvent en contenir jusqu'à 5, voire 6. L'énergie libre de Gibbs de liaison qui résulte de la formation d'un pont disulfure est d'environ - 3,5 kcal/mol (voir Czaplewski et al., 2004). C'est une énergie importante qui contribue, théoriquement, de manière trés significative à la stabilisation de la structure tridimentionnelle native de la protéine. Cependant, les ponts disulfures ne jouent pas un rôle déterminant dans le repliement des protéines, puisque les intermédiaires adoptent des conformations transitoires qui peuvent être éloignées de celles qui mettent en jeu ces ponts disulfures. b. Nombre de ponts Exemple : quel est le nombre de combinaisons permettant de former 3 ponts disulfures à partir de 6 cystéines ?
De manière générale : pour n ponts disulfures et 2n cystéines, il existe [ (2n)! / 2nn! ] combinaisons. |
La distance entre les 2 carbones α de 2 résidus cystéine impliqués dans un pont disulfure est de 4.2 Å à 7.5 Å.
L'entité structurale formé par 2 cystéines reliées par un pont disulfure s'appelle une cystine. La formation d'un pont disulfure est une réaction d'oxydo-réduction réversible : cette réversibilité dépend du potentiel rédox du milieu et du pH.
|
|
c. La réaction de formation d'un pont disulfure Au cours de la réaction d'échange [thiol - disulfure], un anion thiolate R1S- déplace un soufre du pont disulfure R2-S-S-R3.
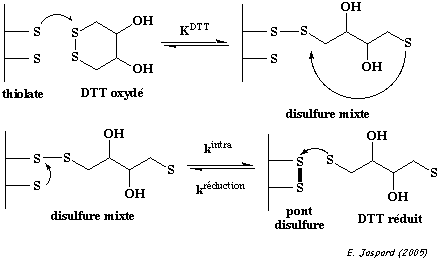
Dans l'état de transition, la charge négative du thiolate est délocalisée sur les 3 atomes de soufre. Les ponts disulfures sont formés et réduits au sein des protéines par 2 réactions d'échange [thiol - disulfure] avec un agent oxydo-réducteur tel que le dithiothréitol (DTT oxydé / DTT réduit) ou le glutathion (GSSG / GSH).
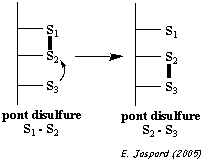
La réaction d'échange [thiol - disulfure] peut aussi avoir lieu au sein d'une même protéine : un groupe thiolate d'une protéine peut attaquer un pont disulfure de la même protéine, conduisant à un remaniement des ponts disulfures.
|
|
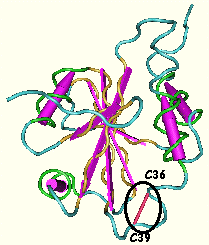
4. Les protéines disulfide isomérases (PDI) et Ero1p Les protéines disulfide isomérases (PDI - EC 5.3.4.1) catalysent le ré-arrangement correct dans le réticulum endoplasmique des ponts disulfures des protéines néo-synthétisées. Ce sont des protéines à plusieurs domaines. Ci-dessous, structure des domaines prolyl 4-hydroxylase de la PDI de Homo sapiens - Kemmink et al. (1996 et 1999).
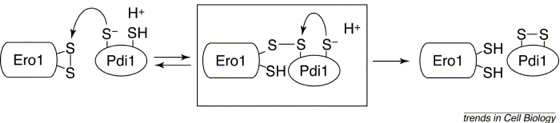
Le site actif du domaine prolyl 4-hydroxylase de la PDI est constitué par les 2 cystéines C36GHC39 qui forment un pont disulfure. Codes : PDB 1MEK et 2BJX Pour oxyder les groupements thiols en disulfure des protéines cibles, les cystéines du site actif de la PDI doivent être maintenues dans l'état oxydé. Ero1p est une flavoenzyme associée à la membrane qui forment des ponts disulfures et les transfère à la PDI. Cette dernière peut alors oxyder les cystéines des protéines substrats. Le mécanisme de l'échange [thiol / disulfure] entre Ero1p et la PDI est le suivant :
Source : Frand et al. (2000)
Source : Gross et al. (2004) Les 5 ponts disulfures intra-chaînes de Ero1p : CYS 90 - CYS 349 / CYS 100 - CYS 105 / CYS 143 - CYS 166 / CYS 150 - CYS 295 / CYS 352 - CYS 355 Ero1p contient 9 sites potentiels de N-glycosylation (Asn-X-Ser/Thr). Le motif de son site actif est : C349XXC352XXC355 |
![]()