| Biogénèse (synthèse, adressage et assemblage) des protéines membranaires intégrales |
| Tweet |
|
|
1. Introduction
2. Rôle clé du réticulum endoplasmique (RE) dans la biogénèse de très nombreuses protéines transmembranaires
3. L'import de protéines précurseurs dans la mitochondrie
|
4. Insertion des protéines de topologie en tonneau β dans les membranes des mitochondries
5. Structure de la membrane interne des mitochondries - Le complexe MICOS 6. Contacts entre le RE et la mitochondrie 7. Biogénèse des protéines des membranes des chloroplastes 8. Biogénèse des protéines des membranes des peroxysomes
9. Les protéines des membranes des bactéries
10. Liens Internet et références bibliographiques |
1. Introduction Toute forme de vie nécessite une barrière physique qui sépare les environnements intracellulaire et extracellulaire : cette barrière est constituée par les différents types de membranes. Environ 25% des gènes codent des protéines membranaires intégrales (PMI). Celles-ci sont insérées :
Les PMI peuvent être classées en :
La structure des PMI répond à deux types d'architecture ou topologie : (i) Le faisceau d'hélices ("α-helix bundle")
(ii) Le tonneau β ("β-barrel")
Les PMI sont essentielles pour de nombreux processus tels que l'adhésion cellulaire et la reconnaissance entre cellules, le trafic membranaire (les divers transports) ou le trafic vésiculaire et leur régulation, les cascades de signalisation, l'établissement de la polarité cellulaire, ... Ces protéines sont donc des cibles thérapeutiques importantes. Leur fonction repose sur leur homo- ou hétéro-oligomérisation et cette complexité, combinée à celles des bicouches lipidiques, rendent particulièrement difficiles leur purification, la détermination de leur stœchiométrie et la cinétique des évènements structuraux (dynamique des complexes membranaires) qui contrôlent leur régulation. Voir un cours sur les constituants des membranes biologiques. |
|
|
| Séquence d'adressage | Courte séquence d'une chaîne polypeptidique qui dirige l'adressage (la destination finale) d'une protéine vers une membrane spécifique. La séquence d'adressage ou peptide signal est située en général à l'extrémité N-terminale ou dans le premier domaine transmembranaire. Elle peut être ou non hydrolysée. Voir un complément sur les peptide signaux. |
| Peptide signal ou séquence signal | Séquence d'adressage qui se trouve à l'extrémité N-terminale des protéines sécrétées et de certaines protéines membranaires. Une fois qu'ils ont rempli leur fonction d'adressage, les peptides signaux sont hydrolysés par une peptidase du signal. |
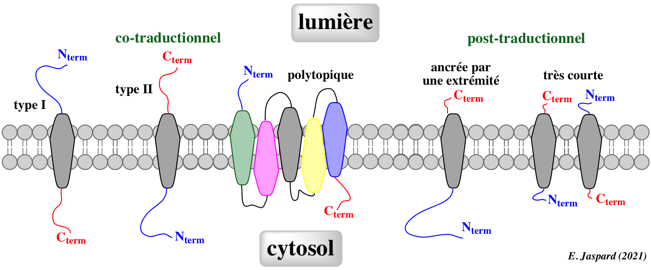
| Protéines membranaires de type I (topologie Nexo) | PMI contenant un peptide signal tel que l'extrémité N-terminale de la chaîne polypeptidique mature est orientée vers la lumière après hydrolyse de ce peptide signal. |
| Protéines membranaires de type II (topologie Ncyt) | PMI dont l'extrémité N-terminale de la chaîne polypeptidique mature est orientée vers le cytosol. |
| Protéines membranaires de type III | PMI dont l'extrémité N-terminale est orientée vers la lumière (topologie Nexo). Ces protéines possèdent une région N-terminale courte (< 50 acides aminés) flanquante. |
| Protéines membranaires ancrées par une extrémité | PMI dont l'unique domaine transmembranaire se situe à environ 65 acides aminés de l'extrémité C-terminale et dont l'extrémité N-terminale est orientée vers le cytosol. Ces protéines sont appelées "tail-anchored (TA) proteins". |
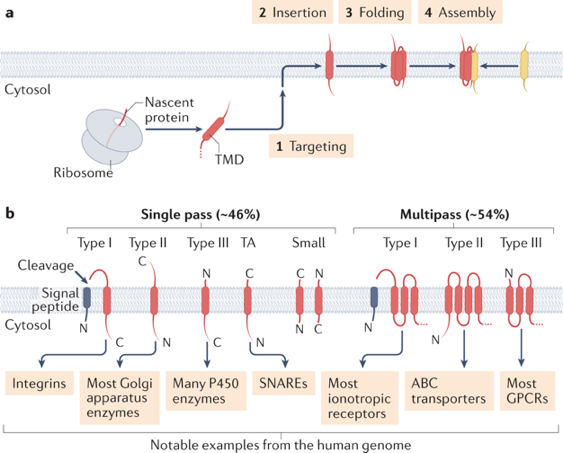
| Protéine membranaire à un passage ou multipasses | Protéine ou sous-unité dont la chaîne polypeptidique traverse la membrane une fois ou plusieurs fois (multipasses). |
| Complexe Sec61 (eucaryotes) ou complexe SecY (procaryotes) | Complexe protéique hétérotrimérique qui transloque des segments polypeptidiques hydrophiles au travers d'une membrane via un canal et insère des domaines hydrophobes dans cette membrane via une porte latérale ("lateral gate"). |
| Insertase | Protéines transmembranaires contenant une poche hydrophile qui facilite la translocation de courts segments polypeptidiques au travers de la membrane en même temps que l'insertion du domaine transmembranaire. |
| Superfamille Oxa1 | Groupe d'insertases liées sur le plan évolutif : il comprend Oxa1 dans la membrane interne de la mitochondrie, YidC dans la membrane interne des bactéries, Ylp1 dans la membrane plasmique des archées, Alb3 dans la membrane interne du chloroplaste, GET1, EMC3 et TMCO1 dans le RE. |
| Chaperon intramembranaire | Facteur du repliement des chaînes polypeptidiques dans la membrane : il protége les domaines transmembranaires (partiellement hydrophiles) des polypeptides naissants jusqu'à leur assemblage avec d'autres domaines transmembranaires. |
| Chaperonine | Famille de complexes multimèriques avec une fonction chaperon dirigée par l'hydrolyse de l'ATP. Leur structure est un cylindre avec une chambre interne qui fournit un environnement protégé dans lequel la chaîne polypeptidique des protéines naissantes se replient. |
| Facteur d'assemblage | Facteur qui favorise l'assemblage de deux ou plusieurs protéines, éventuellement en protégeant temporairement l'interface entre leurs sous-unités. |
| Source : Hegde & Keenan (2021) | |
b. Principes généraux de la biogenèse des protéines transmembranaires C'est l'un des processus cellulaires les plus anciens et ses mécanismes sont remarquablement conservés. La biogenèse des protéines membranaires peut être divisée en 4 étapes :
Source : Hegde & Keenan (2021) Le génome humain code environ 5.000 PMI composées de environ 20.000 domaines transmembranaires qui diffèrent considérablement par leur séquence, par leurs propriétés biophysiques, par leur(s) emplacement(s) dans la protéine et par leur topologie. La machinerie d'adressage et d'insertion doit donc assumer de nombreuses fonctions délicates pour insérer une telle diversité de PMI dans les divers types de membranes.
En conséquence, la machinerie d'insertion doit reconnaître, orienter et fournir un chemin dans la membrane pour une gamme extrêmement diverse de séquences, et donc de structures, protéiques. La plupart des PMI de la cellule peuplent la membrane plasmique et les compartiments subcellulaires des voies de sécrétion et d'endocytose. Ces protéines sont initialement assemblées au niveau du RE, où les domaines transmembranaires des PMI sont intégrés dans la membrane et où la topologie finale est acquise.
|
2. Rôle clé du RE dans la biogénèse de très nombreuses protéines transmembranaires Le RE joue un rôle essentiel dans la synthèse, les modifications et le transport de molécules biochimiques utilisées à l'intérieur et à l'extérieur de la cellule.
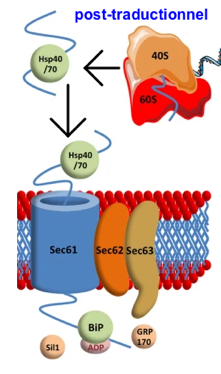
L'import des protéines dans le RE est un processus essentiellement co-traductionnel mais certaines PMI sont insérées dans le RE de manière post-traductionnelle :
|
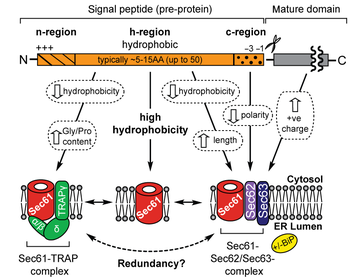
a. Les séquences d'adressage vers le RE et les peptides signaux du RE Günter Blobel a reçu le Prix Nobel en 1999 pour sa découverte que les protéines ont des signaux intrinsèques qui régissent leur transport et leur localisation dans la cellule. Une séquence d'adressage située à au moins 65 acides aminés de l'extrémité C-terminale est requise pour un adressage co-traductionnel.
La plupart des peptides signaux (ou séquences signal) du RE sont constitués :
Source : O'Keefe & High (2020) Les peptides signaux contenant une région h hydrophobe peuvent ouvrir seuls le canal Sec61. Les protéines qui possèdent un peptide signal de faible hydrophobicité nécessitent des composants accessoires :
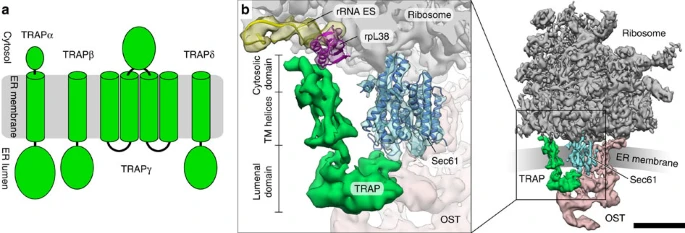
Source : Pfeffer et al. (2017) - OST : oligosaccharyltransferase Sec62/Sec63 facilitent également la translocation co-traductionnelle de protéines sécrétoires avec des peptides signaux caractérisés généralement par une région h plus longue avec une hydrophobicité plus faible et une région c de polarité réduite. |
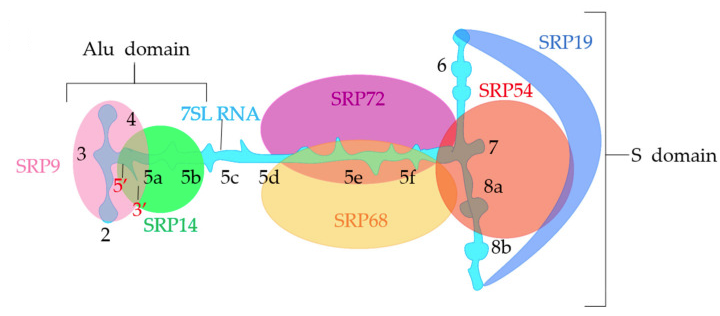
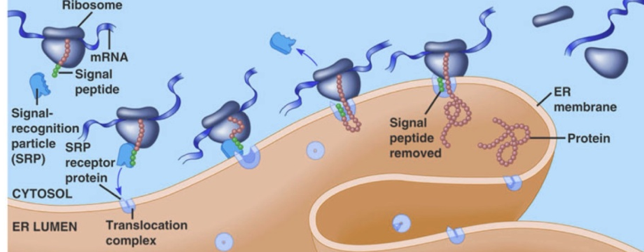
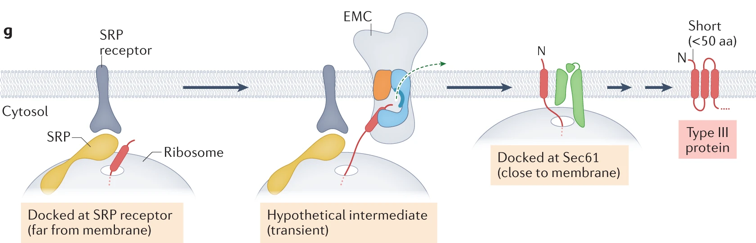
b. La particule de reconnaissance du signal ("Signal Recognition Particle" - SRP) Le complexe SRP, le ribosome et la RNAse P sont les 3 particules ribonucléoprotéiques conservées dans tous les règnes du vivant. Chez les bactéries, SRP est constituée d'une molécule d'ARN 4,5S et de la protéine Ffh (homologue de la sous-unité SRP54, voir ci-dessous). Chez les eucaryotes, le complexe SRP peut être structuralement et fonctionnellement divisé en 2 domaines :
Source : Morgana et al. (2021)
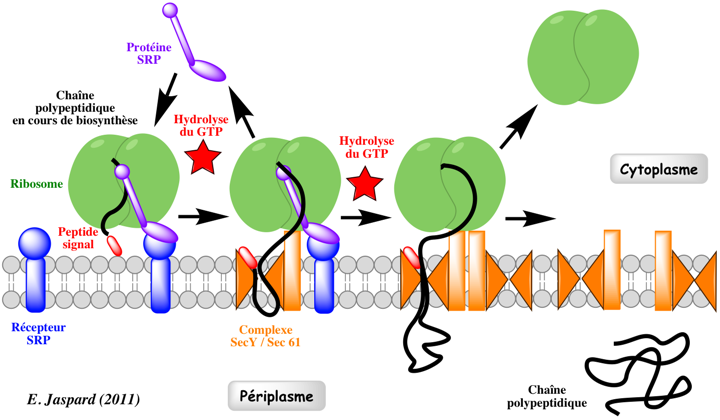
Mécanisme Voir une belle vidéo du mécanisme : "Signal-Recognition-Particle". Le complexe SRP se fixe au récepteur de SRP :
Le complexe SRP ralentit l'élongation de la chaîne polypeptidique en cours de biosynthèse ("elongation arrest"). La chaîne polypeptidique en cours d'élongation emprunte le canal pour passer entièrement dans la lumière du RE (dans le cas d'une protéine sécrétée, dirigée vers l'appareil de Golgi) ou pour être intégrée à la membrane (dans le cas d'une PMI). Il s'agit donc d'un transport co-traductionnel puisque l'élongation de la chaîne polypeptidique continue pendant le passage dans le RE. Cependant, chez les eucaryotes, SRP participe également :
Le complexe SRP et le récepteur de SRP se dissocient pour participer à la translocation d'une autre chaîne polypeptidique en cours de biosynthèse Le cycle SRP est un processus dont l'énergie est fournie par l'hydrolyse du GTP : le complexe SRP et le récepteur de SRP possèdent une activité GTPase.
Source : Numerade L'ARN 7SL
L'ARN 7SL est constitué de 4 domaines (I-IV) composés de 12 hélices (1-12) qui contiennent 4 motifs conservés qui constituent les sites d'interaction avec les sous-unités de SRP. L'ARN 7SL se replie dans une structure secondaire double brin cruciforme où le domaine S en forme de "Y" est la région centrale de l'ARN. Les sous-unités SRP9 et SRP14 possèdent une structure super-secondaire [αβββα], topologie similaire à celle des protéines de liaison à l'ARN double brin : elles forment un hétérodimère stable qui reconnaît le motif "UGUNR" de l'ARN, localisé à l'extrémité 5' hautement conservée du domaine Alu de l'ARN 7SL. Conformément à son rôle dans l'arrêt de l'élongation, le domaine Alu se lie à l'interface avec le ribosome (voir la figure 8 de l'article Soni et al., 2021). Le récepteur de SRP est un hétérodimère :
|
c. Les canaux de translocation ou translocons De nombreuses chaînes polypeptidiques doivent être transportées au travers d'un canal "conducteur" de protéines :
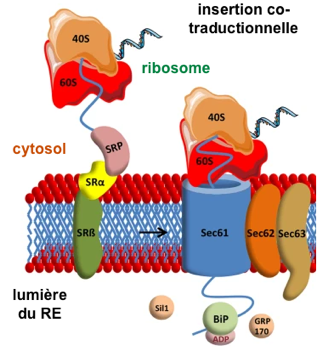
(i) Les protéines qui possèdent une séquence signal hydrophobe sont déplacées de manière co-traductionnelle au travers du canal Sec61 (ou SecYEG) :
40S et 60S : sous-unités 40S et 60S du ribosome; SR : récepteur hétérodimérique de la SRP (ii) Les protéines qui possèdent une séquence signal moins hydrophobes contournent SRP et sont déplacés au travers du canal après la traduction :
Source des deux figures : Linxweiler et al. (2017) BiP-ATPase ("Endoplasmic reticulum chaperone BiP", E.C. 3.6.4.10) :
|
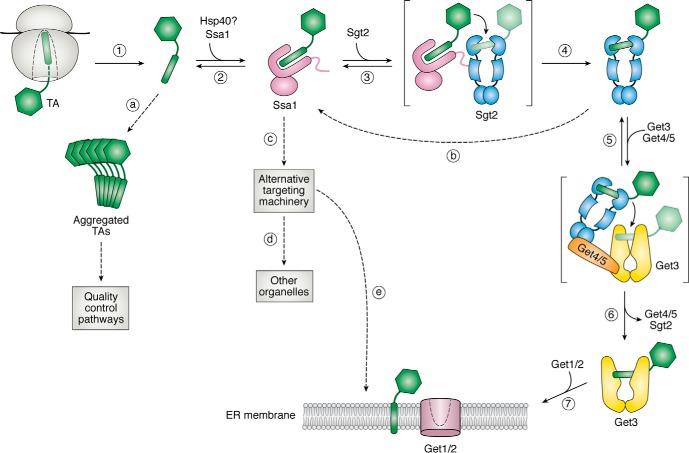
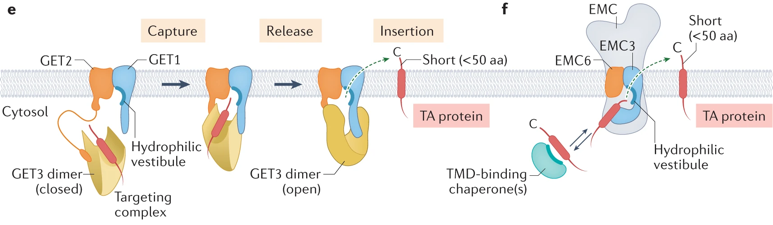
d. La voie GET d'insertion des protéines transmembranaires La voie GET ("Guided-Entry of TA proteins") est requise pour l'insertion des protéines membranaires ancrées par une extrémité (ou "Tail-Anchored (TA) proteins") dans le RE de la levure et des mammifères. Certains gènes orthologues ont également été identifiés chez les plantes supérieures.
Source : Shu-ou Shan (2019)
|
| Composant | Masse molaire |
Caractéristiques |
| GET1 (WRB) | ≈ 20 kDa | Récepteur membranaire (comme GET2) de GET3 qui se fixe de manière sélective au domaine transmembranaire des protéines TA dans le cytosol. Requis pour assurer la topologie correcte de CAMLG et son insertion dans le RE. |
| GET2 (ou CAML ou CAMLG - "Calcium signal-modulating cyclophilin ligand") | ≈ 33 kDa | Récepteur membranaire (comme GET1) de GET3. Nécessaire pour la stabilité de GET1. Stimule la signalisation du calcium dans les cellules T grâce à son implication dans l'élévation du calcium intracellulaire. |
| GET3 (TRC40) | ≈ 39 kDa | ATPase qui reconnaît et se fixe de manière sélective au domaine transmembranaire des protéines TA dans le cytosol. Le complexe [GET3 - protéine TA] cible ensuite le RE (via les récepteurs membranaires GET1 et GET2) où la protéine TA est libérée pour l'insertion. Ce processus est contrôlé par la fixation puis l'hydrolyse de l'ATP : (i) la fixation de l'ATP induit un changement de conformation de l'homodimère GET3 dans un état "fermé" facilitant la reconnaissance des protéines TA nouvellement synthétisées; (ii) l'hydrolyse de l'ATP est nécessaire pour l'insertion. GET3 revient dans l'état "ouvert" ce qui diminue son affinité pour le récepteur GET1 : celui-ci est renvoyé vers le cytosol pour initier un nouveau cycle d'adressage. |
| GET4 (TRC35) | ≈ 36 kDa | Au sein du complexe [GET4/GET5/BAG6 ("BCL2-associated athanogene cochaperone 6")] qui assure le contrôle de qualité du repliement des protéines cytosoliques, GET4 maintient les protéines contenant des régions mal repliés et hydrophobes dans un état "soluble" et participe à leur adressage au RE ou, au contraire, favorise leur envoi vers le protéasome où elles sont dégradées. |
| GET5 (UBL4A) | ≈ 18 kDa | Si le domaine transmembranaire de la protéine TA est fortement hydrophobe, celle-ci est transférée à GET3 via le complexe [GET4-GET5]. GET3 cible ensuite le complexe récepteur [GET1-GET2]. Si le domaine transmembranaire est de faible hydrophobicité, il n'est pas transféré à GET3 : la protéine TA est maintenue dans le cytosol via des interactions avec des chaperons (telles que SGTA, HSP70 et autres). |
| SGTA (Sgt2) | ≈ 34 kDa | Co-chaperone cytoplasmique riche en glutamine qui sert d'échafaudage pour réunir GET4, GET5 et d'autres sous-unités. |
Get3, Get4 et Get5 et Sgt2 sont les homologues chez la levure de TRC40, TRC35, UBL4A et SGTA chez l'homme, respectivement. |
||
e. Le complexe protéique membranaire du RE (EMC) Chez l'homme, le complexe protéique EMC ("ER membrane protein Complex") est constitué de 10 sous-unités (≈ 200 x 70 x 100 Å) qui a une fonction d'insertase membranaire selon un processus indépendant de l'énergie (hydrolyse de l'ATP). L'EMC de la levure contient 8 sous-unités. Le complexe EMC insère :
Schématiquement, la réaction catalysée par le complexe EMC est : protéine ancrée par l'extrémité C-terminale avec des domaines transmembranaires faiblement hydrophobes (associée à la calmoduline dans le cytoplasme) ou PMI de type III => protéine ancrée par l'extrémité C-terminale à la membrane du RE avec des domaines transmembranaires faiblement hydrophobes ou PMI de type III
Voir : "The Endoplasmic Reticulum Membrane Protein Insertion Complex (EMC) Family" - base de données "Transporter Classification Database" |
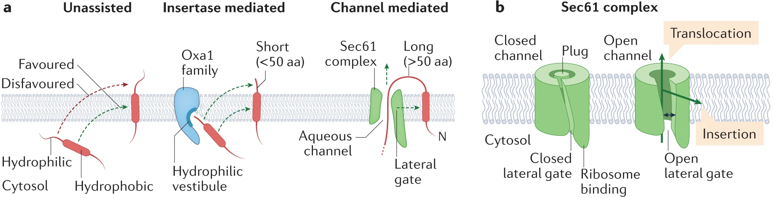
f. Synthèse des voies d'insertion des protéines transmembranaires dans le RE De multiples voies d'insertion fonctionnent en parallèle au niveau du RE pour tenir compte de la grande diversité de structures (topologies) et de propriétés physico-chimiques des PMI à insérer. (a) Comparaison de l'insertion non assistée, médiée par l'insertase et médiée par les canaux pour les domaines transmembranaires (DTM).
(b) Le complexe Sec61 contient un canal aqueux ("aqueous channel") pour la translocation des protéines et une porte latérale ("lateral gate") pour l'insertion de segments hydrophobes dans la membrane. Quand il est fermé, un domaine particulier appelé "plug domain" obture ce canal.
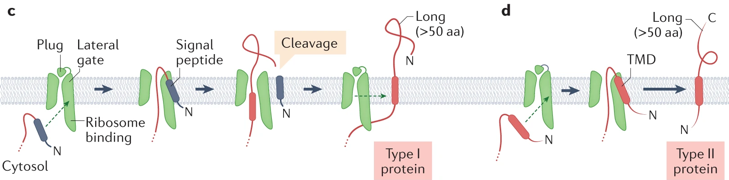
Source : Hegde & Keenan (2021) (c) Une PMI de type I contient un peptide signal qui engage la porte latérale de Sec61. Le segment polypeptidique qui suit est enfilé au travers du canal Sec61 puis le peptide signal est hydrolysé. Un DTM situé en aval pénètre dans le canal Sec61 puis dans la membrane via la porte latérale de Sec61. (d) Une PMI de type II utilise son premier DTM pour engager la porte latérale de Sec61 de la même manière qu'un peptide signal. Le DTM pénètre dans la membrane par la porte latérale et le polypeptide situé en aval est transloqué par le canal Sec61.
Source : Hegde & Keenan (2021) (e) Insertion par la voie d'entrée guidée d'une protéine ancrée par une extrémité ("Guided Entry of Tail-anchored (TA) protein" - GET).
(f) Insertion d'une protéine ancrée par une extrémité par le complexe protéique membranaire du RE ("ER membrane protein complex" - EMC).
Source : Hegde & Keenan (2021) (g) Insertion de PMI de type III par l'EMC.
Source : Hegde & Keenan (2021) |
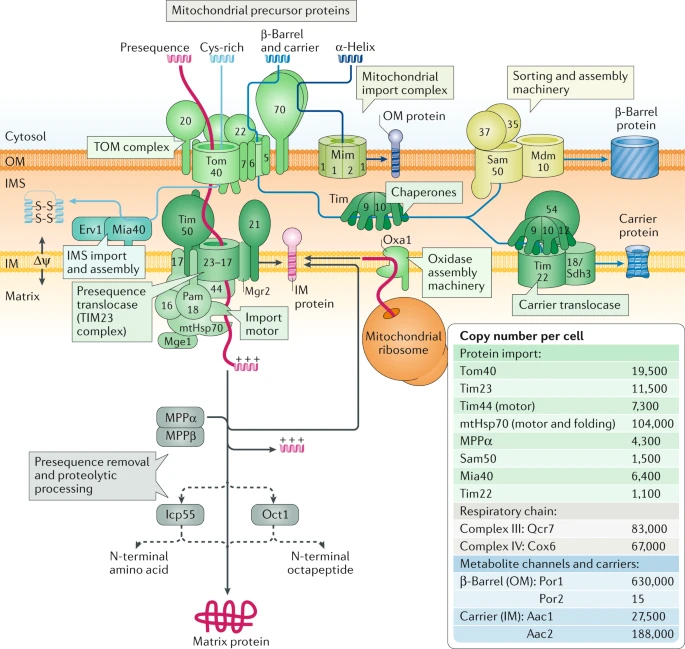
3. L'import de protéines précurseurs dans la mitochondrie Voir un rappel sur la structure, le génome et les propriétés des mitochondries. Du point de vue ultra-structural, la mitochondrie est constituée de 2 membranes (externe et interne) séparées par l'espace inter-membranaire et qui englobent la matrice. Seules 13 protéines mitochondriales sont codées par le génome de la mitochondrie :
En conséquence, plus de 99% des ≈ 1500 protéines mitochondriales sont synthétisées sous forme de précurseurs par les ribosomes dans le cytosol :
|
b. Les grandes voies d'import dans la mitochondrie Différentes voies d'import des pré-protéines du cytosol ont été identifiées selon les organismes :
Le choix du translocon (ou machineries d'import ) dépend de :
Source : Pfanner et al. (2019) α. La voie d'import des protéines qui possèdent une pré-séquence hydrolysable ("the presequence pathway")
Dans les autres voies d'import des protéines dans la mitochondrie, les protéines précurseurs ne contiennent pas de pré-séquences hydrolysables : elles possèdent différents signaux d'adressage. β. La voie de la translocase porteuse ("the carrier pathway") : les précurseurs des protéines porteuses hydrophobes multipasses de la membrane interne sont importés par TOM, par les petits chaperons TIM de l'espace intermembranaire et par la translocase porteuse de la membrane interne ("carrier Translocase of the Inner Membrane" - TIM22). γ. La voie d'import des protéines de topologie en tonneau β ("the β-barrel pathway") : les précurseurs des protéines de la membrane externe de la mitochondrie dont la structure a une topologie en tonneau β utilisent le complexe TOM et des petits chaperons TIM, suivis d'une insertion dans la membrane externe par la machinerie de tri et d'assemblage ("Sorting and Assembly Machinery " - SAM). δ. La voie d'import des protéines qui possèdent un motif spécifique de cystéines : de nombreuses protéines de l'espace intermembranaire mitochondrial sont petites et contiennent des motifs spécifiques de cystéines (formation ou non de ponts disulfures dépendante de l'état rédox). Ces protéines sont importées via TOM et la machinerie d'import et d'assemblage mitochondrial ("Mitochondrial Import and Assembly" - MIA) de l'espace intermembranaire. ε. La voie d'import des protéines qui possèdent des segments transmembranaires α-hélicoïdaux : certaines protéines de la membrane externe qui possèdent des segments transmembranaires α-hélicoïdaux sont importées par le complexe d'import mitochondrial ("Mitochondrial Import" - MIM) : ce complexe permet l'import de protéines qui possèdent une séquence signal d'ancrage N-terminale et l'import de protéines multipasses (polytopiques). |
| Dénomination des composants des machineries d'import des protéines mitochondriales chez la levure | |||
| Nom | Nom chez la levure | Alias | Description - Fonction |
| Complexe translocase de la membrane externe - TOM ("Translocase of the Outer Membrane") | |||
| Tom70 | YNL121C | Mas70, Mom72, Omp1 | Récepteur de protéines précurseurs non hydrolysables |
| Tom40 | YMR203W | Isp42, Mom38 | Pore d'import général |
| Tom22 | YNL131W | Mas17, Mas22, Mom22 | Récepteur central |
| Tom20 | YGR082W | Mas20, Mom19, Pom23, Rir16 | Récepteur de pré-séquence |
| Tom7 | YNL070W | Mom7, Yok22 | Facteur de stabilité / assemblage |
| Tom6 | YOR045W | Isp6, Mom8b | Facteur de stabilité / assemblage |
| Tom5 | YPR133W-A | Mom8a | Facteur de transfert et d'assemblage des pré-protéines |
| Machinerie de tri et d'assemblage de la membrane externe - SAM ("Sorting and Assembly Machinery") | |||
| Sam50 | YNL026W | Omp85, Tob55 | Composant principal conservé du complexe SAM |
| Sam37 | YMR060C | Mas37, Pet3027, Tom37 | Sous-unité du complexe SAM |
| Sam35 | YHR083W | Fmp20, Tob38, Tom38 | Sous-unité du complexe SAM |
| Mdm10 | YAL010C | Fun37 | Coopère avec le complexe SAM et Mdm12/Mmm1 |
| Mdm12 | YOL009C | ---- | Composant de la voie d'assemblage des protéines en tonneau β |
| Mmm1 | YLL006W | Yme6 | Composant de la voie d'assemblage des protéines en tonneau β |
| Mim1 | YOL026C | Tom13 | Impliqué dans la biogenèse des protéines de la membrane externe |
| Chaperons de l'espace intermembranaire ("Chaperones of intermembrane space" - small TIM proteins) | |||
| Tim13 | YGR181W | ---- | Sous-unité du complexe chaperon Tim8-Tim13 |
| Tim10 | YHR005C-A | Mrs11 | Sous-unité du complexe chaperon Tim9-Tim10 et du complexe TIM22 |
| Tim9 | YEL020W-A | ---- | Sous-unité du complexe chaperon Tim9-Tim10 et du complexe TIM22 |
| Tim8 | YJR135W-A | DDP1 | Sous-unité du complexe chaperon Tim8-Tim13 |
| Machinerie d'import et d'assemblage de l'espace intermembranaire - MIA ("Mitochondrial Intermembrane space import and Assembly machinery") | |||
| Mia40 | YKL195W | Tim40, Fmp15 | Récepteur de l'espace intermembranaire de petites protéines précurseurs contenant des cystéines |
| Erv1 | YGR029W | ---- | Sulphydryl oxydase qui fonctionne avec Mia40 |
| Hot13 | YKL084W | ---- | Factor d'assemblage |
| Complexe translocase porteuse de la membrane interne - TIM22 ("Carrier translocase of inner membrane") | |||
| Tim54 | YJL054W | ---- | Sous-unité du complexe TIM22 |
| Tim22 | YDL217C | ---- | Sous-unité centrale du complexe TIM22 qui forme un canal |
| Tim18 | YOR297C | ---- | Sous-unité du complexe TIM22 |
| Tim12 | YBR091C | Mrs5 | Sous-unité périphérique du complexe TIM22 associée à Tim9 et Tim10 |
| Complexe translocase de pré-séquence de la membrane interne - TIM23 ("Presequence translocase of inner membrane") | |||
| Tim50 | YPL063W | ---- | Sous-unité du complexe TIM23 orientée vers l'espace intermembranaire |
| Tim23 | YNR017W | Mas6, Mim23, Mpi3 | Sous-unité du complexe TIM23 qui forme un canal |
| Tim21 | YGR033C | Fmp17 | Interagit avec le complexe TOM et la chaîne respiratoire |
| Tim17 | YJL143W | Mim17, Mpi2, Sms1 | Régulateur du traffic des pré-protéines dans le canal TIM23 |
| Moteur associé à la translocase de pré-séquence - PAM ("Presequence translocase-Associated Motor") | |||
| mtHSP70 | YJR045C | Ens1, Ssc1 | Chaperon central de PAM |
| Tim44 | YIL022W | Isp45, Mim44, Mpi1 | Partenaire de l'HSP70 au niveau de la membrane interne |
| Pam18 | YLR008C | Tim14 | Protéine JDP ("J-domain protein") de la membrane interne complexée à Pam16 |
| Pam17 | YKR065C | Fmp18 | Organisateur du module Pam18 - Pam16 |
| Pam16 | YJL104W | Mia1, Tim16 | Régulateur de Pam18 |
| Mge1 | YOR232W | mtGrpE, Yge1 | Facteur d'échange de nucléotides |
| Source : Bolender et al. (2008) | |||
Dénomination des composants des complexes d'import des protéines mitochondriales chez différents organismes Famille 3.A.8 : "The Mitochondrial Protein Translocase (MPT) Family" (base de données "Transporter Classification Database") |
||||
| Complexe | Levure / Fungi | Homme | Plantes | Trypanosomatidae |
| SAM/TOB | Sam50/Tob55, Sam35/Tob38, Sam37/Mas37, Mdm10 | Sam50/Tob55, metaxine 1, metaxine 2 | Sam50/Tob55, Sam37 (metaxine), Mdm10 | Sam50/Tob55 |
| TOM (translocase de la membrane externe) | Tom40, Tom22, Tom5, Tom6, Tom7, Tom20, Tom70 | Tom40, Tom22, Tom5, Tom6, Tom7, Tom20, Tom70 | Tom40, Tom9 (Tom22), Tom5, Tom6, Tom7, Tom20, OM64 | Atom40, Atom46, Atom69, Atom19, Atom14, Atom11, Atom12 |
TOM est un complexe composé de Tom5, Tom6, Tom7, Tom20, Tom22, Tom40 et Tom70 :
|
||||
| MIM | Mim1, Mim2 | ----- | ----- | Atom36 |
| MIA | Mia40, Erv1 | ----- | Mia40, Erv1 | Erv1 |
| Small Tims | Tim9, Tim10, Tim8, Tim13, Tim12 | Tim9, Tim10a, Tim10b, Tim8, Tim13 | Tim9, Tim10, Tim8, Tim13 | TbTim9, Tbim10, TbTim11, TbTim12, TbTim13, TbTim8/13 |
| TIM23 | Tim23, Tim17, Tim50, Tim21 | TIMM23, TIMM17, TIMM50 | Tim23, Tim17, Tim50, Tim21 | TbTim17, TbTim62 , TbTim54, TbTim42, TbTim50, ACAD, Rhomboid I, Rhomboid II, small TbTims |
| PAM | Pam18, Pam16 (Tim16), Pam17, Tim44, Hsp70, Mge-1 | DnaJC19 et DnaJC15 (homologues de Pam18 de la levure), Magma (TIMM16), mortaline/HSPA9 (Hsp70 mitochondriale) |
Pam18, Pam16, Tim14, Tim44, Hsp70, Mge1 | TbPam27, Hsp70 |
| TIM22 | Tim22, Tim54, Tim18, Sdh3, Tim12 | TIMM22, TIMM29 , AGK | Tim22 | ----- |
| OXA | Oxa1 | Oxa1 ("OXidase Assembly 1-like protein") | Oxa1 | Oxa1 |
OXA est essentielle pour :
|
||||
| Source : Chaudhuri et al. (2021) | ||||
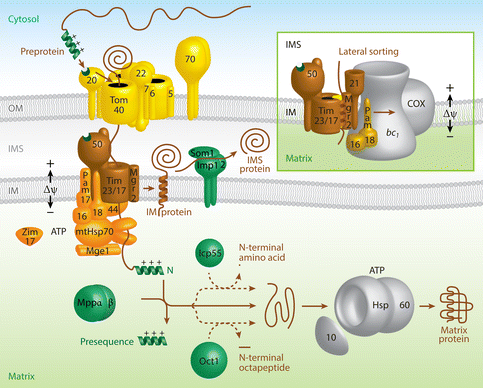
c. La voie d'import des protéines qui possèdent une pré-séquence hydrolysable ("the presequence pathway") C'est la voie d'import vers la matrice mitochondriale : les protéines de la matrice représentent environ 60% des protéines de la mitochondrie.
Source : Wiedemann & Pfanner (2017) Les protéines précurseurs possèdent un signal d'adressage N-terminal appelé pré-séquence qui forme des hélices amphipathiques chargées positivement. Ces pré-séquences sont généralement reconnues par les récepteurs TOM (Tom20 et Tom22) à la surface des mitochondries et elles dirigent la translocation au travers du translocon principal de la membrane externe Tom40.
Le moteur associé à la translocase de pré-séquence ("Presequence translocase-Associated Motor" - PAM) contient l'HSP70 mitochondriale (chaperon central piloté par l'ATP) et 5 cinq co-chaperons. PAM favorise la translocation de l'ensemble de la chaîne polypeptidique dans la matrice. Les pré-séquences sont éliminées par la peptidase hétérodimérique de traitement mitochondrial ("Mitochondrial Processing Peptidase" - MPP - E.C. 3.4.11.21). Des enzymes de la matrice sont impliquées dans le contrôle qualité des chaînes polypeptidiques :
Enfin les protéines sont repliées dans leur forme native - active par l'HSP70 et d'autres protéines chaperons telles que HSP10 - HSP60. |
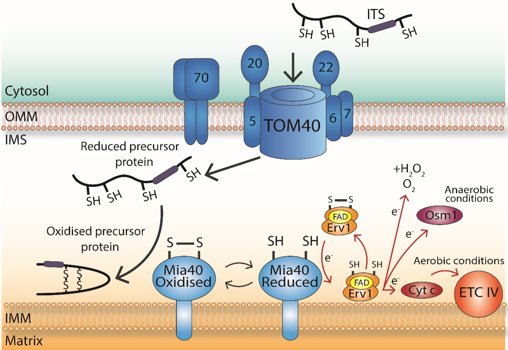
d. La voie d'import des protéines qui possèdent un motif spécifique de cystéines Certaines petites protéines ["small cysteine-containing proteins (small Tim)" - 8 à 22 kDa], codées par le génome nucléaire, qui contiennent un motif double CX3-9C sont d'abord transloquées par la tranlocase TOM de la membrane externe. Puis la machinerie d'import et d'assemblage mitochondrial ("Mitochondrial Import and Assembly" - MIA) de l'espace intermembranaire crée des ponts disulfure au sein des protéines transloquées ce qui les maintient dans l'espace intermembranaire des mitochondries.
Source : Dickson-Murray et al. (2021) La séquence signal hydrophobe ITS ("Intermembrane space Targeting Signal") interagit avec Mia40 qui contient 6 cystéines au sein d'un motif CPC-CX9C-CX9C :
|
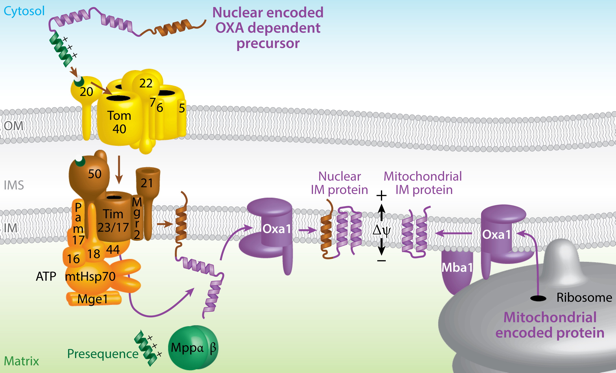
e. OXA1 : la voie d'import/export de la matrice vers la membrane interne de la mitochondrie OXA1 ("Oxidase Assembly 1") a plusieurs rôles :
Source : Wiedemann & Pfanner (2017) |
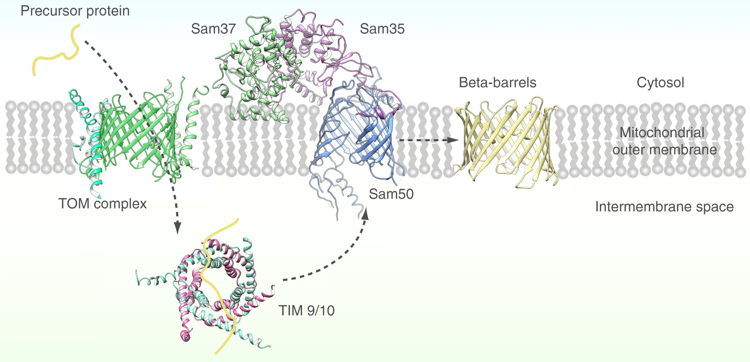
4. Insertion des protéines de topologie en tonneau β ("β-barrel") dans les membranes des mitochondries La membrane externe de la mitochondrie contient des protéines qui ont une topologie structurale en tonneau β ("β-barrel proteins") qui établissent une communication entre le cytosol et l'intérieur de la mitochondrie. L'insertion de protéines en tonneau β dans la membrane externe est médiée par la machinerie de tri et d'assemblage mitochondrial multi-sous-unités SAM("multisubunit mitochondrial Sorting and Assembly Machinery", également appelée TOB).
Source : Diederichs et al. (2020) On trouve également une topologie structurale en tonneaux β dans :
|
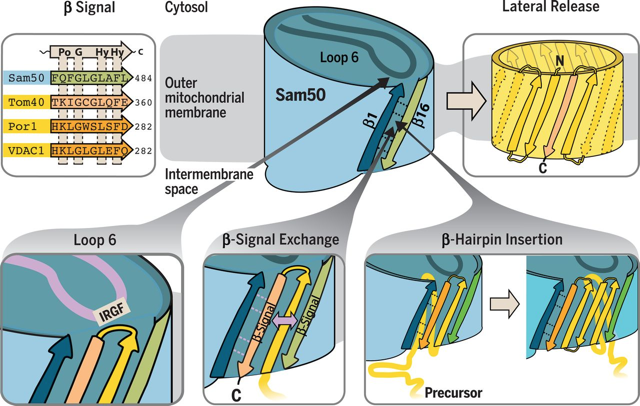
b. Rôle de la porte latérale de Sam50 dans l'insertion des protéines en tonneau β Le précurseur de la protéine en tonneau β est transféré via Sam50 jusqu'à la porte latérale formée par le brin β 1 et le brin β 16. Lors de l'ouverture de la porte latérale, le signal de la protéine précurseur remplace le signal β endogène de Sam50. Une boucle conservée dans Sam50 ("loop 6" - motif de liaison IRGF) favorise la fixation de la séquence signal à la porte latérale puis l'insertion des structures β dites en épingles à cheveux ("β hairpins") qui suivent. La protéine en tonneau β repliée est libérée dans la membrane externe.
Source : Höhr et al. (2018) Caractéristiques de Sam50 Sam50 (ou Tob55) est un membre de la superfamille Omp85 typique des membranes externes des bactéries Gram-, des mitochondries et des chloroplastes. La caractéristique de cette superfamille est son domaine transmembranaire C-terminal en tonneau β.
|
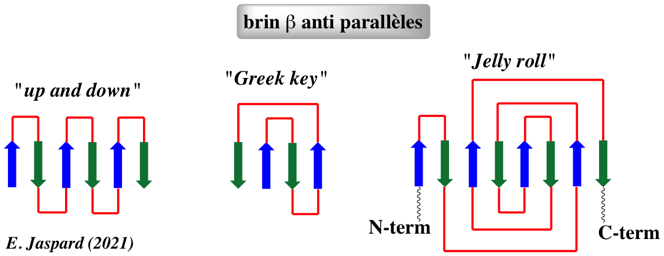
c. Caractéristiques des protéines en tonneau β La topologie en tonneau β est caractérisée par le nombre n de brins antiparallèles et par le nombre S de cisaillements ("shear") qui mesure l'angle d'inclinaison (36° à 44°) des brins par rapport à l'axe du tonneau. Exemples de motifs et de repliements en tonneau β :
Le domaine "small β-barrel" :
Le repliement "double-psi β barrel" :
Le repliement OB-fold ("Oligonucleotide/Oligosaccharide-Binding fold)" :
|
Visualisation de Tom40 de de Neurospora crassa à une résolution de 6,8 Å Structure déterminée par cryomicroscopie électronique. Code PDB : 5O8O
|
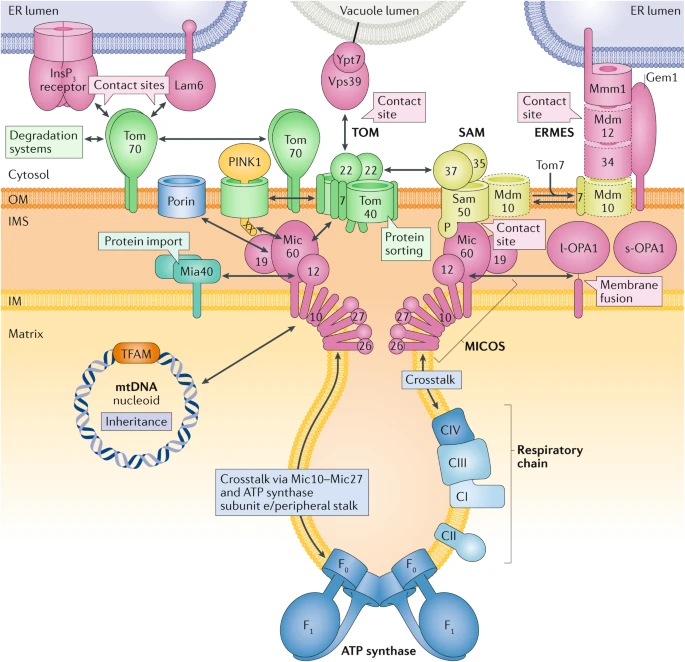
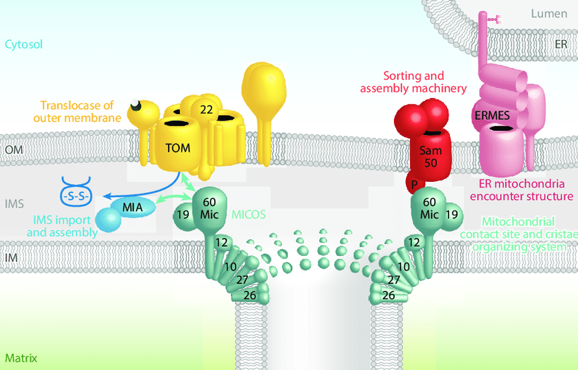
5. Structure de la membrane interne des mitochondries - Le complexe MICOS La partie de la membrane interne des mitochondries qui est parallèle à la membrane externe est appelée membrane interne limite ou bordante ("inner boundary membrane"). L'autre partie de la membrane interne de la mitochondrie forme des crêtes de dimensions variables vers la matrices : elle est appelée membrane des crêtes ("cristae membrane"). Cette structure lui confèrent une très grande surface où ont lieu les réactions biochimiques. Les mitochondries dont les crêtes ont une structure incorrecte sont associées à de très graves pathologies. La formation, la structure, le remodelage dynamique et les jonctions des crêtes résultent de l'action de divers acteurs : le complexe appelé système d'organisation des crêtes et des sites de contact mitochondriaux ("MItochondrial contact site and Cristae Organizing System" - MICOS), la GTPase OPA1 de type dynamine, l'ATP synthase F1-F0 (courbure positive de la membrane) et la cardiolipine.
Source : Pfanner et al. (2019) Le complexe MICOS est composé de sept sous-unités (Mic10, Mic13, Mic19, Mic25, Mic26, Mic27 et Mic60) :
MICOS interagit avec SAM via le domaine associé au transport de polypeptides ("the POlypeptide TRAnsport-associated (POTRA) domain") de Sam50. |
6. Contacts entre le RE et la mitochondrie a. Le complexe ERMES ("ER - Mitochondria Encounter Structure") C'est un complexe protéique composé de 4 protéines qui relie le RE et ala mitochondrie :
b. Le réseau d'organisation ERMIONE ("endoplasmic reticulum (ER) - MItochondria Organizing NEtwork") entre la mitochondrie et le RE Les sites de contacts entre les membranes externe et interne de la mitochondrie et les sites de contacts entre la membrane externe de la mitochondrie et le RE sont des éléments cruciaux d'un vaste réseau qui contrôle la biogenèse des protéines et des lipides, l'architecture et la dynamique des membranes, le transport des ions et des métabolites, la dynamique mitochondriale et l'hérédité.
Source : Wiedemann & Pfanner (2017) En effet, le complexe MICOS de la membrane interne de la mitochondrie, la translocase TOM de la membrane externe de la mitochondrie et la machinerie SAM de la membrane externe de la mitochondrie forme le coeur du grand réseau d'organisation ERMIONE. Ce réseau établit une multitude d'interactions dynamiques avec :
En conclusion, ERMIONE forme un système transmembranaire qui coordonne la biogenèse des protéines et des lipides, les aspects énergétiques, l'hérédité et le contrôle qualité des mitochondries (biogénèse et morphologie). |
7. Biogénèse des protéines des membranes des chloroplastes Environ 95% des protéines chloroplastiques sont codées par le génome nucléaire et doivent être importées dans les chloroplastes :
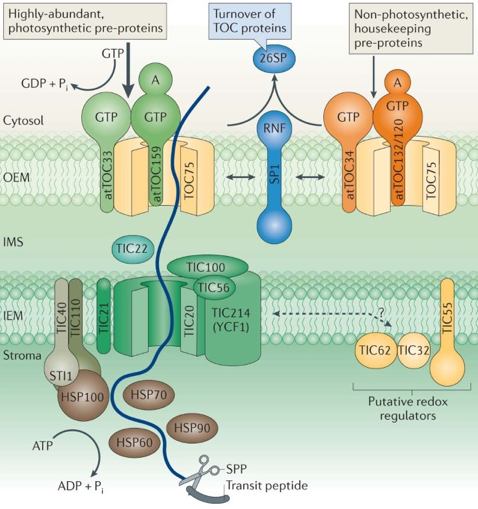
L'import dans les plastes des protéines précurseurs traduites dans le cytosol est réalisé par les translocons de l'enveloppe externe et de l'enveloppe interne du chloroplaste ("Translocon at the Outer/Inner envelope of the Chloroplast"), TOC et TIC respectivement, et des chaperons solubles.
a. Translocation au travers de la membrane externe des chloroplastes Le complexe TOC (enveloppe externe) pour la translocation de précurseurs photosynthétiques est composé de Toc33, Toc64, Toc75 et Toc159. Le complexe TOC pour l'import de précurseurs non photosynthétiques est composé de Toc34, Toc75, Toc120 et Toc132.
La protéine précurseur est d'abord transloquée par le complexe TOC. Si elle est destinée à être intégrée dans la membrane externe sous forme de tonneau β, elle est transférée à Oep80 ("Chloroplast Outer Envelope Protein 80 kDa", TCDB 1.B.33.2.2) qui la replie et l'intègre dans cette membrane.
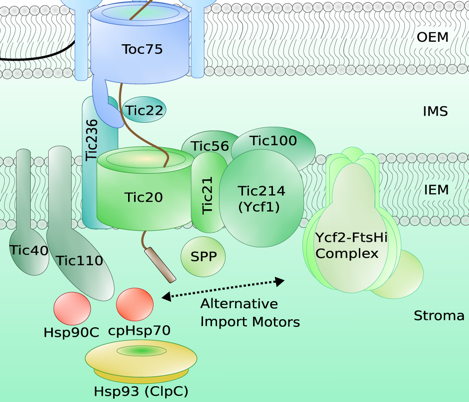
Source : Jarvis & Lopez-Juez (2013) b. Translocation au travers de la membrane interne des chloroplastes La translocation au travers de la membrane interne est médiée par le complexe TIC composé de Tic20, Tic21, Tic22, Tic56, Tic100 et Tic214.
Source : Thomson et al. (2020) Le peptide d'adressage est traité en 2 étapes :
|
8. Biogénèse des protéines des membranes des peroxysomes Les peroxysomes sont des organites à 1 membrane présents dans presque toutes les cellules eucaryotes. Les fonctions des peroxysomes englobent notamment :
Le contenu, le nombre et la taille des peroxysomes peuvent changer rapidement en réponse aux signaux environnementaux. Les peroxysomes superflus et/ou endommagés par l'oxydation sont dégradés par un processus appelé pexophagie. La biogenèse des peroxysomes implique au moins 37 protéines appelées peroxines (codées par les gènes Pex) qui ont un rôle dans divers processus :
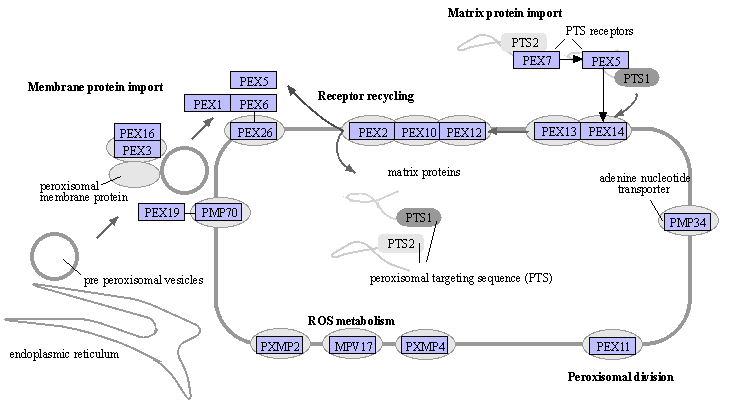
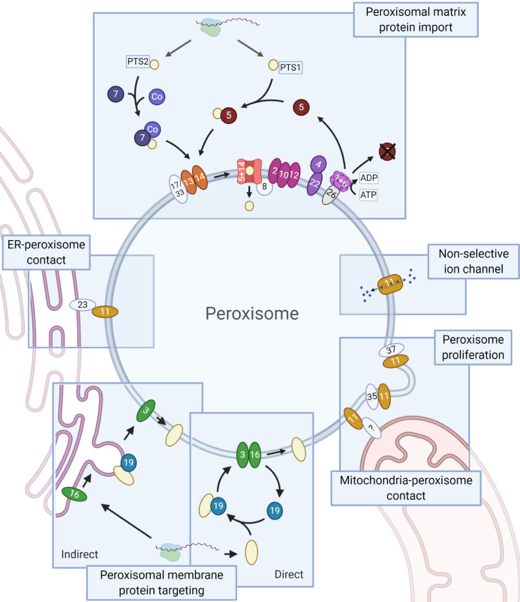
Source : KEGG (La figure du site KEGG est interactive et permet d'accéder à un trés grand nombre d'informations) |
b. Mécanismes d'import dans la membrane et dans la matrice Les peroxysomes ne possèdent ni génome ni machinerie de synthèse protéique : ils doivent importer leurs protéines (après traduction dans le cytosol) dans leur membrane. Deux voies d'adressage et d'insertion co-existent. α. La voie de tri direct Pex19 se fixe aux PMP nouvellement traduites dans le cytosol et agit comme protéine chaperon en interagissant avec leurs domaines hydrophobes couvrant la membrane. Le complexe [Pex19-PMP] se lie à la peroxine membranaire intégrale Pex3 et est inséré dans la membrane par un mécanisme encore inconnu. β. Le transit intermédiare via le RE La détection de PMP à l'intérieur du RE est à l'origine d'un modèle suggèrant que le RE est à l'origine de la biogenèse des peroxysomes en médiant le trafic des PMP vers les peroxysomes via des vésicules bourgeonnantes.
Source : Jansen et al. (2021) Import des protéines dans la matrice peroxysomale Les peroxysomes doivent également importer les protéines dans leur lumière appelée matrice peroxysomale. Ce mécanisme importe des protéines entièrement repliées (voire sous formes d'oligomères). Schématiquement, l'import dans la matrice des peroxysomes est catalysée par une dizaine de PMP intégrales ou périphériques qui : (i) amarrent le complexe [récepteur d'import / protéine à transloquer] à la membrane peroxysomale; (ii) transloquent la protéine dans la matrice de l'organite par un pore de translocation; (iii) exportent le récepteur vers le cytosol et le recyclent. Après synthèse dans les ribosomes, les protéines qui contiennent les signaux d'adressage aux peroxysomes ("Peroxisomal Targeting Signals") PTS1 (extrémité C-terminale) ou PTS2 (extrémité N-terminale) se fixent à leur récepteur respectif, Pex5 ("Peroxisomal Targeting Signal 1 Receptor") ou Pex7, et forment un complexe [récepteur- protéine à transloquer].
Le recyclage du récepteur implique :
|
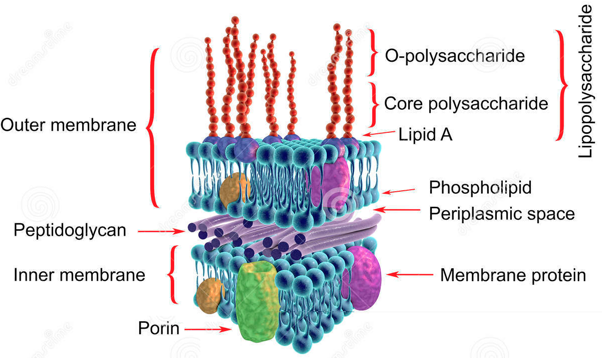
9. Les protéines des membranes des bactéries Les bactéries Gram- ont une enveloppe cellulaire qui comprend une membrane interne, une couche de peptidoglycane et une membrane externe.
Source : Dreamstime La membrane externe des bactéries à Gram- est constituée d'un feuillet interne de phospholipides et d'un feuillet externe de lipopolysaccharides réticulés par des cations divalents chélatés. Cette membrane asymétrique forme une structure robuste qui stabilise l'enveloppe cellulaire. La membrane externe est un facteur de virulence des bactéries pathogènes. C'est également une composante majeure de la résistance aux antibiotiques des bactéries Gram- :
Les récepteurs TonB (PF00593) de la membrane externe des bactéries Gram- forment une famille de protéines en tonneau β :
|
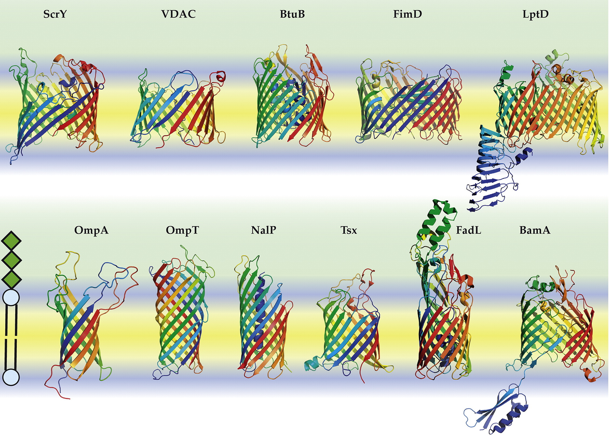
b. Biogénèse des protéines en tonneau β de la membrane externe Les protéines de la membrane externe ("Outer Membrane Proteins" - OMPs) des bactéries forment des tonneaux β transmembranaires avec un nombre pair (8 à 36) de brins et 8 à 24 cisaillements. Environ 2,5% des gènes des bactéries Gram- codent des porines en tonneau β. Les toxines bactériennes comme l'hémolysine adoptent également une topologie en tonneau β. Exemples de protéines bactériennes en tonneau β :
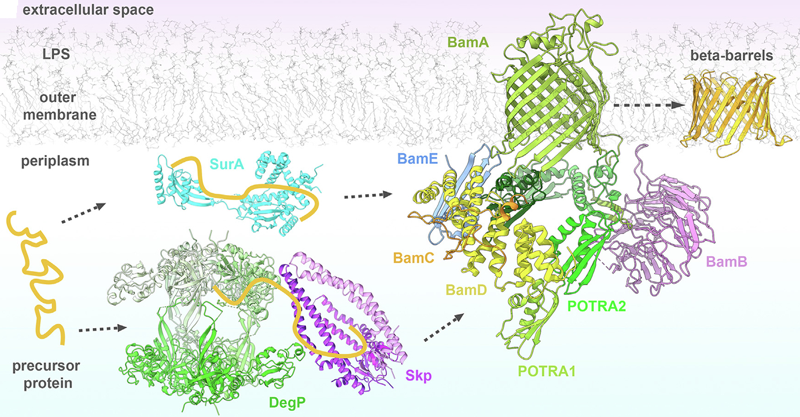
Source : Kleinschmidt J.H. (2015) Les protéines en tonneau β sont synthétisées dans le cytosol puis sécrétées dans le périplasme au travers de la membrane interne par la translocase SEC. Enfin, l'assemblage et l'intégration des protéines en tonneau β dans la membrane externe des bactéries Gram- est catalysée par la machinerie d'assemblage des protéines en tonneau β ("Bacterial β-barrel Assembly Machinery" - BAM) :
Source : Diederichs et al. (2021) |
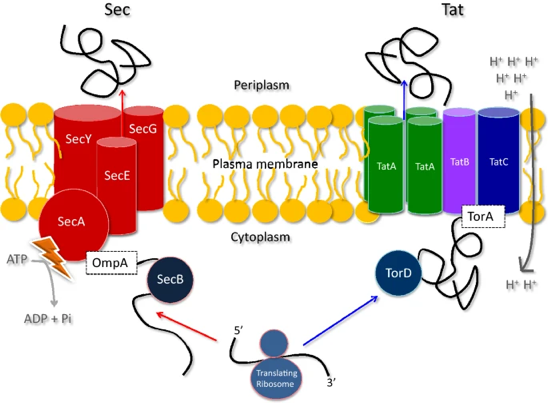
c. Protéines de la membrane interne des bactéries : voies SEC et TAT La membrane interne des bactéries Gram- est une bicouche phospholipidique d'environ 6 nm d'épaisseur. Elle forme une barrière semi-perméable, entre le cytoplasme et le périplasme, qui contrôle l'export et l'import d'ions, de polymères d'oses, d'ADN et de protéines. La modélisation suggère que la fixation d'OmpA ("Outer membrane protein A" - membrane externe) et de TolR ("Tol-Pal system protein TolR" - membrane interne - environ 900 molécules par cellule d'Escherichia coli) au peptidoglycane maintient la position de la paroi cellulaire dans le périplasme approximativement à égale distance des membranes interne et externe. Il semble que la protéine trimérique Lpp ("Major outer membrane Lipoprotein") contrôle la largeur du périplasme, ajuste son angle d'inclinaison pour s'adapter à l'espace disponible et peut compenser en partie l'absence d'OmpA. La voie sécrétoire SEC
La voie de translocation d'arginines jumelles ("Twin-Arginine Translocation" - TAT)
Source : Frain et al. (2019) Figure ci-dessus :
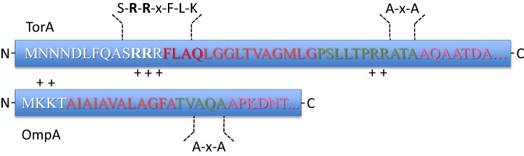
Les peptides signaux des voies TAT et SEC Le peptide signal N-terminal des chaînes polypeptidiques naissantes les dirige vers la translocase correcte SEC ou TAT.
Source : Frain et al. (2019) Par ailleurs, les peptides signaux TAT sont :
|
| 10. Liens Internet et références bibliographiques |
|
"Endoplasmic Reticulum, Golgi Apparatus, and Lysosomes" (Scitable Nature Education) "Tertiary Protein Structure and Folds" |
|
|
Castillo et al. (1999) "A six-stranded double-psi β barrel is shared by several protein superfamilies" Structure 7, 227 - 236 G. von Heijne (2007) "The membrane protein universe: what's out there and why bother ?" J. Intern. Med. 261, 543 - 557 Bolender et al. (2008) "Multiple pathways for sorting mitochondrial precursor proteins" EMBO reports 9, 42 - 49 Fairman et al. (2011) "The structural biology of β-barrel membrane proteins: a summary of recent reports" Curr. Opin. Struct. Biol. 21, 523 - 531 Jarvis & Lopez-Juez (2013) "Biogenesis and homeostasis of chloroplasts and other plastids" Nat. Rev. Mol. Cell Biol. 14, 787 - 802 |
|
|
Cymer et al. (2015) "Mechanisms of integral membrane protein insertion and folding" J. Mol. Biol. 427, 999 - 1022 Höhr et al. (2015) "Assembly of β-barrel proteins in the mitochondrial outer membrane" Biochim. Biophys. Acta 1853, 74 - 88 Kleinschmidt J.H. (2015) "Folding of β-barrel membrane proteins in lipid bilayers — Unassisted and assisted folding and insertion" Biochim. Biophys. Acta - Biomembranes 1848, 1927 - 1943 Mayerhofer P. U. (2016) "Targeting and insertion of peroxisomal membrane proteins: ER trafficking versus direct delivery to peroxisomes" Biochim. Biophys. Acta 1863, 870 - 880 |
|
|
Pfeffer et al. (2017) "Dissecting the molecular organization of the translocon-associated protein complex" Nat. Commun. 8, 14516 Linxweiler et al. (2017) "Let's talk about Secs: Sec61, Sec62 and Sec63 in signal transduction, oncology and personalized medicine" Signal Transduct Target Ther. 2, 17002 Wiedemann & Pfanner (2017) "Mitochondrial machineries for protein import and assembly" Annu. Rev. Biochem. 86, 685 - 714 Chaturvedi et al. (2017) "Transmembrane β-barrels: Evolution, folding and energetics" Biochim. Biophys. Acta Biomembr. 1859, 2467 - 2482 Höhr et al. (2018) "Membrane protein insertion through a mitochondrial β-barrel gate" Science 359, eaah6834 |
|
|
Wu et al. (2019) "Structure of the post-translational protein translocation machinery of the ER membrane" Nature 566, 136 - 139 Shu-ou Shan (2019) "Guiding tail-anchored membrane proteins to the endoplasmic reticulum in a chaperone cascade" J. Biol. Chem. 294, 16577 - 16586 Pfanner et al. (2019) "Mitochondrial protein organization: from biogenesis to networks and function" Nat. Rev. Mol. Cell Biol. 20, 267 - 284 Youkharibache et al. (2019) "The small β-barrel domain: a survey-based structural analysis" Structure 27, 6 - 26 Frain et al. (2019) "Transport of folded proteins by the Tat system" Protein J. 38, 377 - 388 |
|
|
O'Keefe & High (2020) "Membrane translocation at the ER: with a little help from my friends" FEBS Journal 287, 4607 - 4611 Pleiner et al. (2020) "Structural basis for membrane insertion by the human ER membrane protein complex" Science 369, 433 - 436 Diederichs et al. (2020) "Structural insight into mitochondrial β-barrel outer membrane protein biogenesis" Nat. Commun. 11, 3290 Thomson et al. (2020) "Protein import into chloroplasts and its regulation by the ubiquitin-proteasome system" Biochem. Soc. Trans. 48, 71 - 82 |
|
|
Hegde & Keenan (2021) "The mechanisms of integral membrane protein biogenesis" Nat. Rev. Mol Cell Biol. doi: 10.1038/s41580-021-00413-2 Morgana et al. (2021) "SRPassing co-translational targeting: the role of the signal recognition particle in protein targeting and mRNA protection" Int. J. Mol. Sci. 22, 6284 Soni et al. (2021) "Structural analysis of the SRP Alu domain from Plasmodium falciparum reveals a non-canonical open conformation" Commun. Biol. 4, 600 Zhang et al. (2021) "Ribosome-bound Get4/5 facilitates the capture of tail-anchored proteins by Sgt2 in yeast" Nat. Commun. 12, 782 |
|
Chaudhuri et al. (2021) "Diverse Functions of Tim50, a Component of the Mitochondrial Inner Membrane Protein Translocase" Int. J. Mol. Sci. 22, 7779 Diederichs et al. (2021) "Building Better Barrels – β-barrel Biogenesis and Insertion in Bacteria and Mitochondria" J. Mol. Biol. 433, 166894 Dickson-Murray et al. (2021) "The Mia40/CHCHD4 oxidative folding system : redox regulation and signaling in the mitochondrial intermembrane space" Antioxidants (Basel) 10, 592 Gaussmann et al. (2021) "Membrane interactions of the peroxisomal proteins PEX5 and PEX14" Front. Cell Dev. Biol. 9, 651449 |
|
Jansen et al. (2021) "Comparative genomics of peroxisome biogenesis proteins: making sense of the PEX proteins" Front. Cell Dev. Biol. 9, 654163 Mendez et al. (2022) "Cryo-EM of CcsBA reveals the basis for cytochrome c biogenesis and heme transport" Nat. Chem. Biol. 18, 101 - 108 Smalinskaite & Hegde (2023) "The Biogenesis of Multipass Membrane Proteins" Cold Spring Harb. Perspect. Biol. 15, a041251 |
![]()