| La glutamine synthétase |
| Tweet |
|
|
1. Formation du glutamate par la glutamate déshydrogénase (E.C. 1.4.1.4) 2. Formation de la glutamine à partir du glutamate par la glutamine synthétase (E.C. 6.3.1.2) 3. Le couple glutamine synthétase - GOGAT (EC 1.4.1.13) |
4. Régulation de l'activité de la glutamine synthétase
5. La glutamine synthétase des plantes 6. Liens internet et références |
|
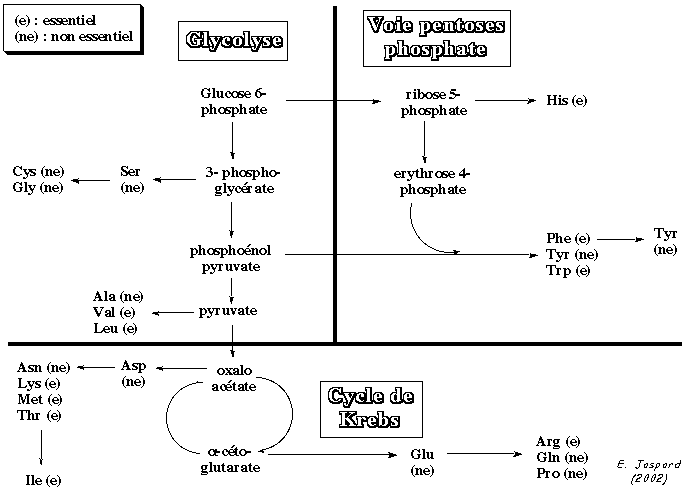
20 acides aminés (les plus courants - on en recense plus de 350) constituent les protéines. On pourrait s'attendre à un grand nombre de voies de biosynthèse de ces composés. La cellule a adopté une stratégie qui repose sur des mécanismes communs à plusieurs acides aminés : ainsi, l'on a des branchements à partir d'un intermédiaire commun. Par ailleurs, les acides aminés ont pour précurseurs des métabolites issus des grandes voies métaboliques comme : la glycolyse, le cycle des acides tricarboxyliques, la voie des pentoses phosphate.
(e) : acides aminés essentiels - (ne) : acides aminés non essentiels |
|
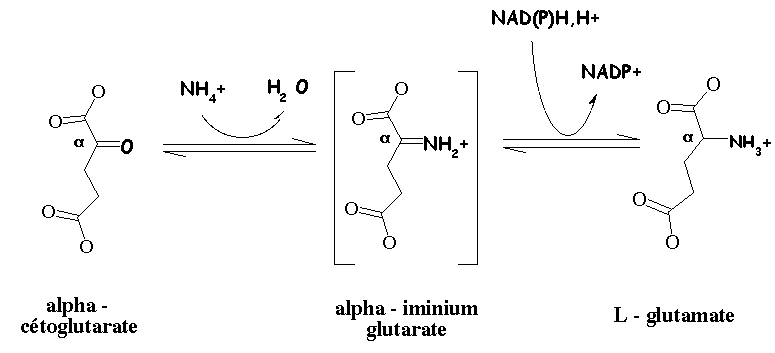
1. Formation du glutamate par la glutamate déshydrogénase E.C. 1.4.1.4 On peut considérer l'assimilation de NH4+ par la glutamate déshydrogénase E.C. 1.4.1.4 (GDH4) comme un point d'entrée dans la synthèse des acides aminés.
Selon les organismes, cette réaction est catalysée principalement dans un sens ou dans l'autre et utilise une coenzyme particulière :
Voir la structure hypothétique de la glutamate déshydrogénase EC 1.4.1.4 |
|
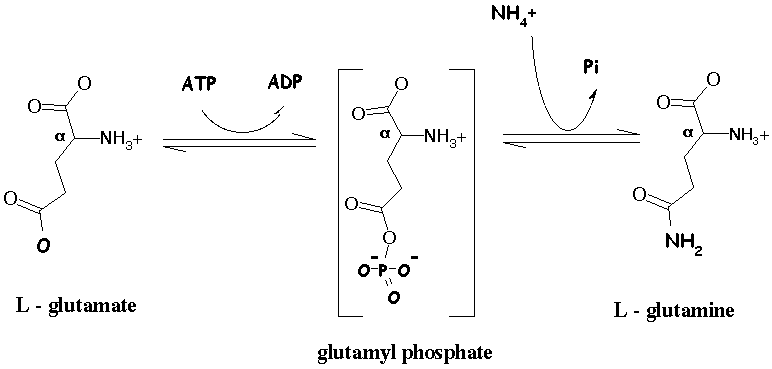
2. Formation de la glutamine à partir du glutamate par la glutamine synthétase La seconde réaction clé dans l'assimilation d'ammoniac correspond à la formation de glutamine à partir de glutamate et d'ammoniac, réaction catalysée par la glutamine synthétase (GS - EC 6.3.1.2).
|
|
3. Le couple glutamine synthétase - GOGAT GOGAT (EC 1.4.1.13) = [NADPH-dependent glutamine:2-oxoglutarate amidotransferase] ou glutamate synthase. Les données expérimentales, obtenues notamment chez les plantes, sont en faveur d'une synthèse du glutamate, non pas par la glutamate déshydrogénase EC 1.4.1.4, mais par le couple GS - GOGAT :
Voir une simulation de l'activité du couple GS - GOGAT : "Computer Simulation of Metabolism Consideration of Cycles - The GS/GOGAT Cycle" |
|
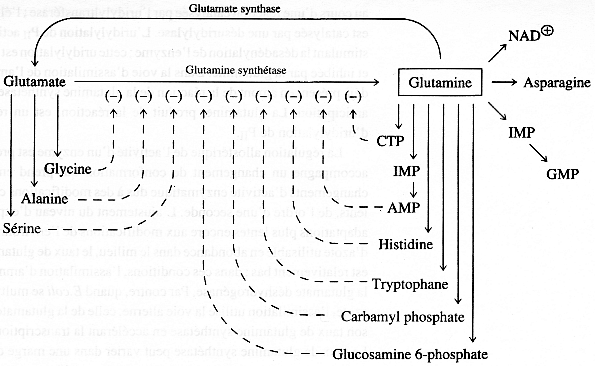
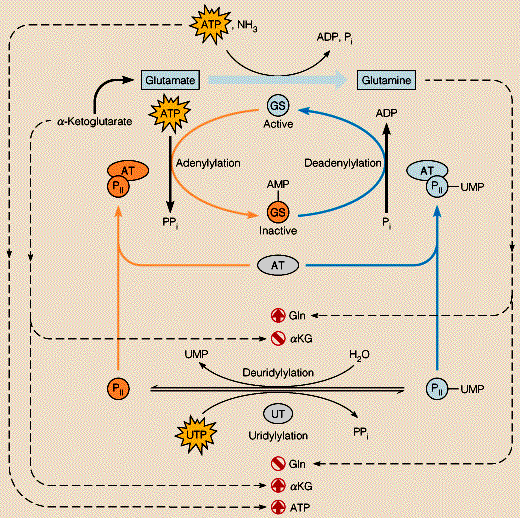
4. Régulation de l'activité de la glutamine synthétase 4a. Rétro - inhibition cumulative de la glutamine synthétase Escherichia coli ne possède qu'une forme de GS codée par le gène glnA. La GS de Escherichia coli est un excellent exemple pour souligner la régulation à laquelle sont soumises la plupart des enzymes qui interviennent dans la synthèse des acides aminés. En effet cette enzyme est un oligomère de 12 sous-unités identiques qui porte chacune non seulement un site actif mais également des sites de fixation d'inhibiteurs allostériques :
Source : "Principe de Biochimie" Horton et al. (1994) Ed. DeBoeck Université On comprend plus aisément l'extrême sophistication de la GS en regard des divers rôles de la glutamine au sein de la cellule :
|
4b. Régulation de l'activité de la glutamine synthétase : adénylyltransférase, protéine PII et uridylyltransférase Chez un grand nombre de bactéries, l'activité de la GS est finement régulée par une adénylyltransférase ou ATase. L'ATase (EC 2.7.7.42) catalyse la réaction : 12 ATP + GS <=> 12 PPi + GS-(AMP)12
L'ATase existe sous 2 formes : l'ATase A (AT en orange dans le shéma ci-dessous) qui adénylyle la GS et l'ATase D qui la désadénylyle (AT en bleu) :
Source : "Regulation of the activity of E. coli glutamine synthetase"
|
| isoforme GS | localisation | fonction |
| GS1 | feuile et tige | synthèse de la glutamine pour le transport d'azote |
| racine | assimilation directe de l'ammoniac ou des nitrates du sol | |
| cotylédon | ré-assimilation de l'ammoniac relarguées par la dégradation des réserves azotées au cours de la germination | |
| GS2 | chloroplaste | assimilation de l'ammoniac produit à partir des nitrates et ré-assimilation de l'ammoniac relarguées par la photorespiration |
| GSN | nodules de racines | assimilation de l'ammoniac sécrété dans le cytosol des plantes inféctées par les bactéries fixatrice d'azote |
|
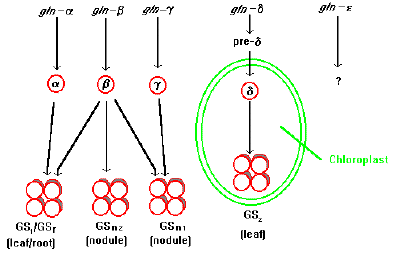
La famille des gènes codant la GS chez les plantes est complexe. L'isoforme de GS des feuilles de Phaseolus vulgaris résulte de l'expression différentielle de 3 membres de cette famille multigènique (figure ci-dessous - Lea et al, 1989) :
|
|
6. Liens internet et références Site web : "Ammonia Assimilation and Recycling - Regulation of glutamine synthetase - Bacteria and Cyanobacteria" Berlett et al. (1998) "Carbon dioxide stimulates peroxynitrite-mediated nitration of tyrosine residues and inhibits oxidation of methionine residues of glutamine synthetase: both modifications mimic effects of adenylylation" Proc. Natl. Acad. Sci. U.S.A. 95, 2784-2789 Caban & Ginsburg (1976) "Glutamine synthetase adenylyltransferase from Escherichia coli: purification and physical and chemical properties" Biochemistry 15, 1569-1580 Stewart et al. (1980) "Enzymes of glutamate formation: glutamate dehydrogenase, glutamine synthetase, and glutamate synthase". In BJ Miflin ed "The Biochemistry of Plants", Vol 5, Academic Press, New York, 271-327 Baima et al. (1989) "Characterization of a cDNA clone for barley leaf glutamine synthetase" Carlsberg Res. Commun. 54, 1-9 Cock et al. (1991) "A nuclear gene with many introns encoding ammonium-inducible chloroplastic NADP-specific glutamate dehydrogenase(s) in Chlorella sorokiniana" Plant Mol. Biol. 17, 1023-1044 Lea et al. (1989) "The use of mutants lacking glutamine synthetase and glutamate synthase to study their role in plant nitrogen metabolism". In JE Poulton, JT Romeo, EE Conn eds "Plant Nitrogen Metabolism", Recent Advances in Phytochemistry Vol 23. Plenum Press, New York, 157-189 |
![]()