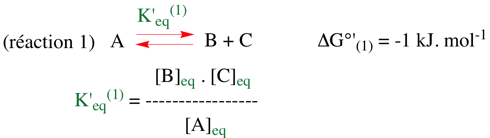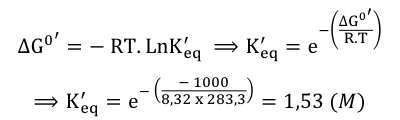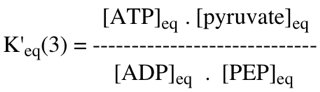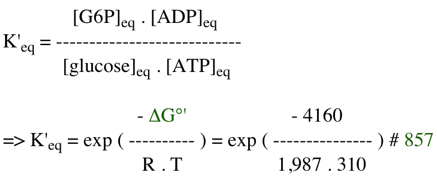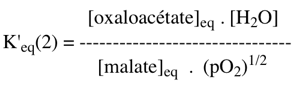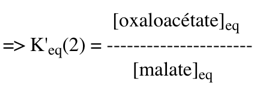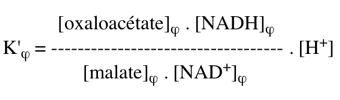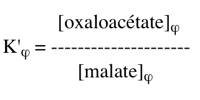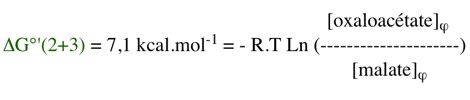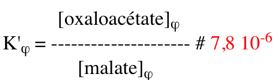| Correction TD Bioénergétique - Equilibre des réactions biochimiques et variation d'énergie libre de Gibbs |
| Tweet |
|
| a. Rappel sur
la variation d'énergie libre de Gibbs dans les conditions standard : ΔG0'
Pour déterminer la variation d'énergie libre de Gibbs d'une réaction dans les conditions standard :
Le sigle 0 traduit que la réaction est effectuée dans des conditions standard et le sigle prime indique que la réaction a lieu à pH 7 (considéré comme le pH physiologique).
b. Rappel sur la variation d'énergie libre de Gibbs dans les conditions cellulaires : ΔG' On mesure les concentrations des métabolites dans la cellule puis on calcule ΔG' (voir ci-dessous). Les conditions n'étant pas standard il n'y a pas le sigle 0 mais il y a le sigle prime car les mesures sont effectuées dans la cellule (pH = 7).
|
Exercice N°1 1. Le clivage d'un composé A donne les produits B et C. La réaction est caractérisée par une variation d'énergie libre de Gibbs dans les conditions standard à pH 7 : ΔG0' = -1 kJ.mol-1.
Très important :
Il faut être très vigilant aux valeurs employées pour ce calcul :
2. On couple la formation de A (à partir de B et C) à l'isomérisation d'un composé D en E. Cette isomérisation est également caractérisée par une variation d'énergie libre de Gibbs dans les conditions standard à pH 7 : ΔG0' = -1 kJ.mol-1.
=> ΔG0'(globale) = - R.T LnK'eq(globale) => K'eq(globale) = 1 (M-1) |
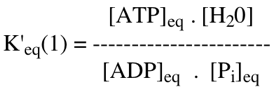
Exercice N°2 1. La réaction de formation de l'ATP s'écrit : ADP + Pi <=> ATP + H2O Cette réaction de la glycolyse est catalysée par la pyruvate kinase. Elle permet la synthèse nette d'ATP au cours de cette voie métabolique.
Remarque : La concentration de l'eau vaut [masse de 1 L d'eau / massse molaire de l'eau] = 1000/18 = 55,5 M.
2. La réaction d'hydrolyse du PEP s'écrit : PEP + H2O <=> pyruvate + Pi
3. La réaction résultant du couplage s'écrit : ADP + Pi + PEP + H2O <=> ATP + H2O + pyruvate + Pi Soit : ADP + PEP <=>ATP + pyruvate
=> K'eq(1) . K'eq(2) = 4,432 10-6 x 7,135 1010 = 3,16 105 => ΔG0'(3) = - RT Ln K'eq(3) = - 7,47 kcal.mol-1 |
Exercice N°3 glucose + ATP <=> glucose-6-phosphate (G6P) + ADP
La constante K'φ traduit le rapport des concentrations physiologiques :
K'eq et K'φ ont des valeurs différentes :
Pour connaître le sens spontané de cette réaction in vivo il faut calculer : ΔG' = ΔG0' + R.T Ln K'φ = - 4160 + [1,987 x 310 x Ln (1,24 10-3)] = - 8,28 kcal.mol-1 Conclusions
|
Exercice N°4 1. La constante d'équilibre de la réaction (2) s'écrit :
Comme : ΔG0'(2) = - RT Ln K'eq(2) avec ΔG0'(2) = - 45300 cal.mol-1 => K'eq(2) = 1033 2. La réaction couplée (2+3) s'écrit : malate + NAD+ <=> oxaloacétate + NADH + H+ C'est une réaction du cycle de Krebs. K'φ est la constante définie par le rapport des concentrations physiologiques des métabolites :
La constante K'φ s'écrit alors :
Par ailleurs : ΔG0'(2+3) = ΔG0'(2) + ΔG0'(3) = (- 45,3 + 52,4) = 7,1 kcal.mol-1
Puisque : ΔG'(2+3) = 0, alors ΔG0'(2+3) = - RT Ln K'φ
La valeur du rapport des concentrations [oxaloacétate]φ / [malate]φ pour que la variation d'énergie libre du couplage soit nulle est donc :
|
Exercice N°5 Comme dans l'exercice 4, ne pas confondre variation d'énergie libre de Gibbs dans les conditions standard (ΔG0') et variation d'énergie libre de Gibbs (ΔG'). Ces 2 grandeurs sont liées par la relation : [B]exp ΔG' = ΔG0' + RT Ln( ---------- ) [A]exp [B]eq Comme : ΔG0' = - RT Ln K'eq = - RT Ln( -------- ) [A]eq [B]eq [B]exp => ΔG' = - RT Ln( --------- ) + RT Ln( ---------- ) [A]eq [A]exp Pour que la réaction soit spontanée, il faut que ΔG'< 0 :
[B]exp [B]eq => RT Ln ( --------- ) < RT Ln( --------- ) [A]exp [A]eq [B]exp [B]eq => ---------- < --------- [A]exp [A]eq [B]exp => --------- < Keq [A]exp |
Exercice N°6 1. La réaction catalysée par la pyruvate kinase s'écrit : PEP + ADP <=> pyruvate + ATP Le PEP est hydrolysé en intermédiare énolate avec libération d'un Pi immédiatement transféré à l'ADP pour former l'ATP. 2. La variation d'énergie libre de Gibbs dans les conditions standard et dans les conditions physiologiques sont respectivement : ΔG0' = - 7500 cal.mol-1 et ΔG' = - 4000 cal.mol-1 (voir l'énoncé).
|
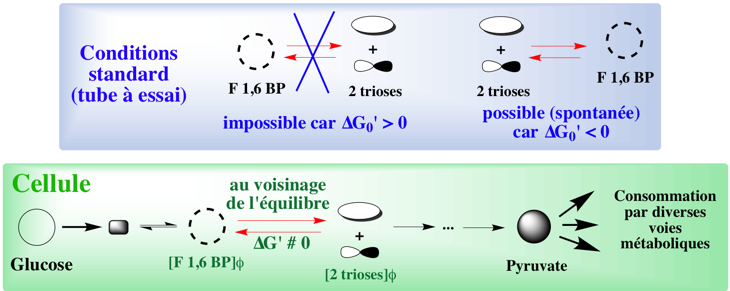
Exercice N°7 La réaction de formation du G3P et de la DHAP s'écrit : fructose 1,6 diP <=> G3P + DHAP 1. Dans les conditions standard, cette réaction est caractérisée par ΔG0' = + 5,79 kcal.mol-1. La constante d'équilibre de cette réaction s'écrit :
Conclusion
|
![]()