
| L'électrophorèse des protéines et des acides nucléiques |
| Tweet |
|
|
1. Principe 2. Gel d'électrophorèse des protéines en conditions dénaturantes 3. Détermination de la masse molaire d'une protéine 4. Gel d'électrophorèse d'ADN |
5. Révélation des bandes d'ADN (coloration) par le bromure d'éthidium 6. Détermination de la taille d'un fragment d'ADN 7. Expérience à analyser 8. Gels bidimensionnel en protéomique |
|
1. Principe L'électrophorèse a pour but de séparer des molécules chargées au travers d'un gel (un polymère) sous l'effet d'un champ électrique. Les molécules se déplacent vers le pôle de charge opposée à leur charge nette z, à une vitesse v proportionnelle à cette charge : v = E. z / f où E est la force du champ éléctrique et f la force de friction. La mobilité électrophorétique d'une molécule peut-être exprimée (de manière simplifiée) par la relation : mobilité = k . (pH - pI) / masse molaire où pI est le point isoélectrique de la molécule. Cette technique permet, entre autre, de :
|
|
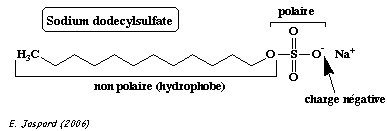
On fait bouillir un mélange de protéines en présence :
En conséquence :
|
|
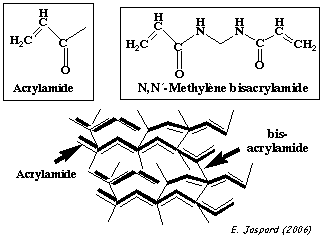
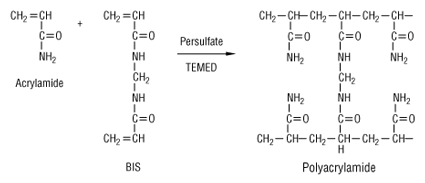
Les protéines de l'échantillon sont ensuite séparées par une électrophorèse sur gel de polyacrylamide en présence de SDS. C'est un gel réticulé, obtenu par polymérisation :
La réaction de polymérisation est :
|
|
|
Plus le pourcentage d'acrylamide est élevé, plus la densité des chaînes est élevée et les mailles du réseau sont serrées. En conséquence :
|
|
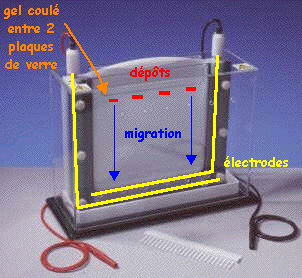
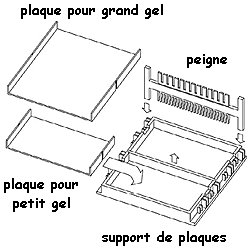
Le gel est coulé entre des plaques de verre fixées sur un support et un peigne est enchâssé entre ces plaques.
Source : Bio-Rad Après polymérisation du gel, le peigne est retiré formant ainsi des puits. La taille et le nombre des dents des peignes sont variables ce qui permet de déposer des volumes allant de 20 µL à 200µL d'échantillon de protéines à séparer. Les plaques de verre contenant le gel polymérisé sont placées dans une cuve d'électrophorèse. Du tampon d'électrophorèse conducteur (électrolytes) est mis dans la cuve et la migration s'effectue sous l'action d'un champ électrique :
Les échantillons de protéines dénaturées sont déposés dans les puits. |
| Après migration, le gel est
démoulé.
Les protéines sont fixées dans le gel par une solution qui contient du méthanol et de l'acide acétique qui dénaturent de manière irréversible les protéines dans les mailles du gel. Les protéines sont révélées par une coloration : par exemple avec le bleu de Coomassie ou le nitrate d'argent (plus sensible).
|
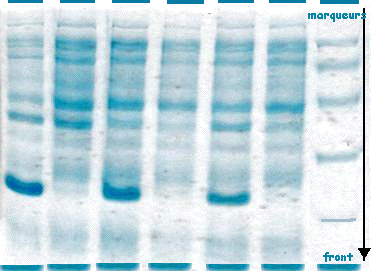
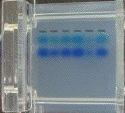
| On obtient
différentes bandes pour chaque piste de la figure
ci-contre (la flèche
indique le sens de migration).
La masse molaire des protéines est déterminée à l'aide de marqueurs qui sont des protéines standards de masses molaires connues (piste de droite). Exemple de marqueurs :
|
Source : E. Jaspard (2004) |
| 4. Gel d'électrophorèse d'ADN |
|
Source : Pharmacia® |
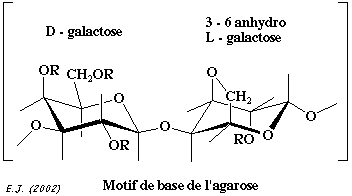
L'agarose (figure ci-contre) est un polymère d'un diholoside (masse molaire d'environ 120.000 Da). |
|
| Description du système i-MUPID® |
|
|
|
|
|
|
Tampon d'électrophorèse
Tampon de charge : bleu de bromophénol 0,02% - xylène cyanol 0,02% - glycérol 3% - tampon TBE. Les 2 colorants permettent de suivre la migration des fragments d'ADN :
|
|
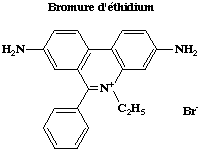
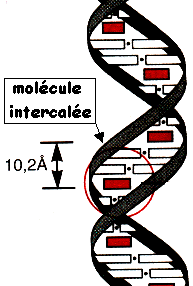
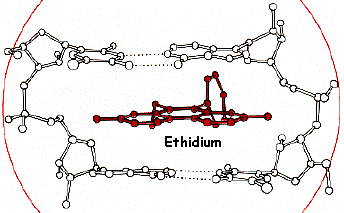
5. Révélation des bandes d'ADN (coloration) par le bromure d'éthidium Des hétérocycles organiques à noyaux accolés comme le bromure d'éthidium se lient à l'ADN bicaténaire par intercalation. Ils occupent un site interbase sur deux jusqu'à saturation. Ce type de liaison est dit par "exclusion du voisinage". Les intercalants sont séparés les uns des autres par des intervalles de 10,2 Å. Le bromure d'éthidium (concentration d'utilisation finale : 0.5 µg/ml) est extrèmement dangereux et nécessite de porter des gants pour manipuler les gels lors de la révélation et après. |
|
|
Intercalation ---> |
|
Source des figures : "Principes de Biochimie Minérale" Lippard & Berg (1997), Ed. DeBoeck Universités |
|
La visualisation des bandes révélées avec le bromure d'éthidium nécessite un transilluminateur UV (longueur d'onde : 302 nm, voire 254 nm et 365 nm). Les rayons UV sont dangereux pour les yeux : il faut porter des lunettes, ou disposer d'une enceinte dans laquelle sont placés le transilluminateur et une caméra. |
|
|
|
|
Il existe des méthodes de coloration de l'ADN qui ne nécessitent pas l'emploi d'UV mais sont moins sensibles que la révélation par le bromure d'éthidium :
La coloration au bleu de Nile semble une méthode à la fois sensible et non dangereuse. |
|
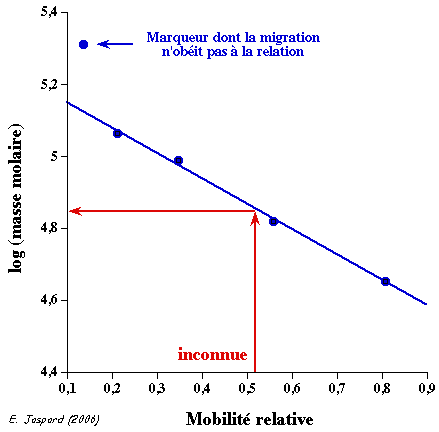
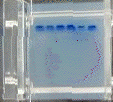
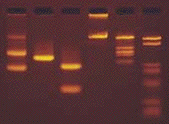
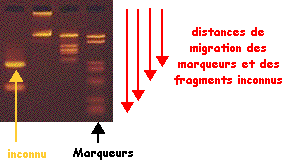
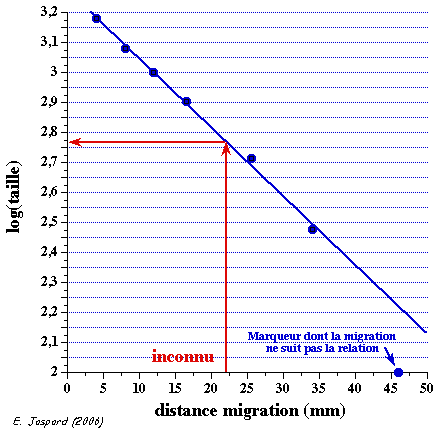
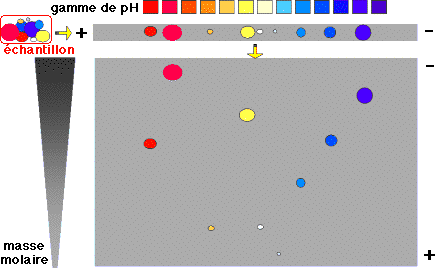
6. Détermination de la taille d'un fragment d'ADN Gel d'électrophorèse sur agarose (ci-dessous) sur lequel on voit :
La droite : log (taille) = f(distance de migration) permet de déterminer la taille en paires de base d'un fragment d'ADN inconnu (figure de droite).
|
|
Voir une simulation de migration électrophorètique d'ADN. Analyse du gel de la figure ci-contre :
|
|
8. L'électrophorèse sur gel bi-dimensionnel en protéomique (SWISS-2DPAGE) a. Principe La stratégie la plus courante est de séparer dans un premier temps les protéines par électrophorèse bidimensionnelle en gel de polyacrylamide (ou 2D - PAGE - "two-dimensional polyacrylamide gel electrophoresis"). En effet, du fait de leur composition en acides aminés, les protéines diffèrent les unes des autres par leur masse molaire et leur charge nette à un pH donné. Cette valeur est caractérisée par le point isoélectrique ou pI. Les protéines ont :
L'une des difficultés majeures de la séparation est de trouver des conditions d'électrophorèse qui couvrent de telles gammes de propriétés physico-chimiques. |
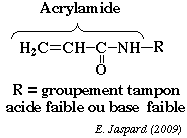
La séparation se fait en deux temps. 1ère dimension : une isoélectrofocalisation (IEF) où les polypeptides migrent jusqu'à un pH égal à leur pl. La séparation selon la charge utilisait auparavant des ampholytes pour obtenir un gradient de pH. Mais ce procédé n'était pas assez reproductible pour la comparaison de gels par superposition. On utilise maintenant des gradients de pH immobilisés (IPG), formés avec des immobilines. Les immobilines sont des dérivés de l'acrylamide et leur structure générale est décrite dans la figure ci-contre. R correspond à un groupement carboxyle ou à une amine tertiaire qui forment une série de molécules tampon avec différentes valeurs de pKa d'ionisation (exemples : 3,6 - 4,4 - 4,6 - 6,2 - 7,0 - 8,5 - 9,3). |
|
Les immobilines co-polymérisent avec les molécules d'acrylamide (voir figure ci-dessus) dans le gel d'IEF. Ainsi, les groupes chargés portés par les immobilines qui forment le gradient de pH sont covalamment attachés (via une liaison vinyle) à la matrice de polyacrylamide et donc immobilisés. Cette technique permet d'obtenir des gradients de pH reproductibles et très étroits : une pente de 0.01 unité pH / cm peut séparer un mélange de protéines avec une différence de pI de 0.001 unité pH ! |
| 2ème dimension : une séparation selon la masse molaire est un gel d'électrophorèse de polyacrylamide en conditions dénaturantes (sodium dodécyl sulfate ou SDS et réduction des ponts disulfure). |
Source : Piercenet |
| Ces deux électrophorèses
sont effectuées de manière perpendiculaire l'une par rapport à l'autre.
Comme les paramètres de la séparation sont indépendants, cette technique est particulièrement résolutive : plusieurs centaines de chaînes polypeptidiques peuvent être séparées sous forme de taches de protéines ou spots sur un gel.
Source : Humboldt - Universität Berlin Voir des exemples de gels bi-dimensionnels. Remarque : de plus en plus de techniques de protéomiques emploient une séparation (ou plusieurs successivement) par la chromatographie liquide à haute performance ("high performance liquid chromatography" - HPLC). Exemples : nano-LC-MS/MS ou 2D-LC-MS/MS. |
b. Les méthodes de révélation des protéines Coloration des gels pour la révélation des protéines (100 ng à 0,1 ng de protéine par spot) :
Source : Sigma- Aldrich Les logiciels d'analyse d'image des gels repèrent les spots et calculent leur coordonnées et leur intensité. La sélection des spots à prélever est transmise au couteau à gel. Les morceaux de gels sont déposés dans les puits de plaques à 96 puits. Exemple d'appareillage :
Voir un développement de la protéomique et de la spectromètrie de masse. |
![]()







{kind=link}