Optimisation des conditions d'expression et purification partielle de la forme mature de la protéine de choc thermique HSP22 recombinante de pois (Pisum sativum) Discussion |
|
Voir un cours sur les protéines de stress LEA et HSP. |
1. Optimisation des conditions d'expression Les souches se distinguent par le taux de base de transcription en absence d'induction. La différence d'intensité relative pour la bande correspondant à la forme mature de l'HSP22 recombinante visualisée dans le cas de BL21(DE3) et dans le cas de B834(DE3)pLysS indique que la "fuite" est nettement moindre en présence du plasmide pLysS. Les conditions d'induction retenues dans ce travail ont été une température de culture des bactéries de 37°C et une concentration en IPTG de 0,4 mM.
Une ambiguïté subsiste : quel est l'effet d'une température susceptible d'induire un choc thermique des bactéries ou, en d'autres termes, à quelle protéine correspond la bande visualisée à 42°C en absence d'IPTG ? |
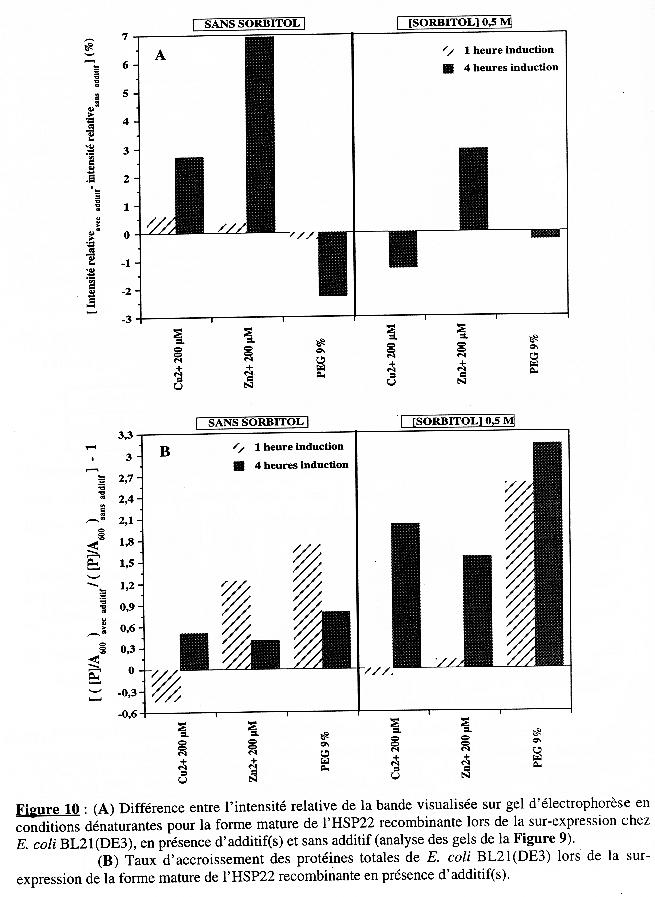
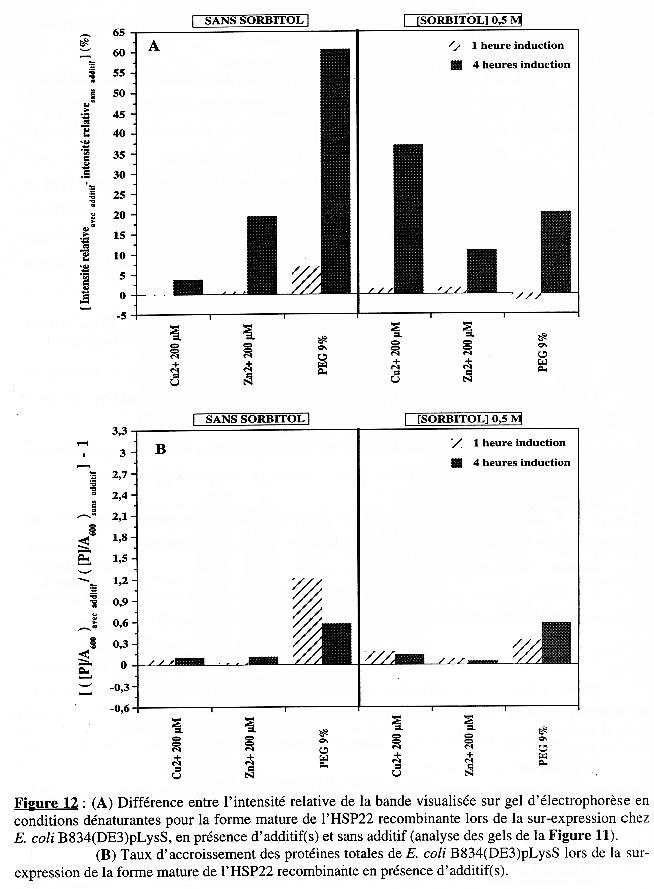
La présence de cuivre et surtout de zinc augmente la sur-expression de la protéine recombinante chez BL21(DE3) mais cette augmentation est moindre en présence de sorbitol. Le taux d'accroissement en protéines totales augmente avec le temps d'induction en présence des 3 additifs et la présence du sorbitol augmente cet effet. Dans le cas de BL21(DE3), l'augmentation de la quantité de protéines observée en présence de zinc et de cuivre (dans une moindre mesure), en absence de sorbitol, est due essentiellement à la synthèse de la forme mature de l'HSP22 recombinante (Figures 10A & 10B). Le zinc n'a pas d'effet sur la synthèse spécifique de la forme mature de l'HSP22 recombinante dans le cas de B834(DE3)pLysS. Quant au taux d'accroissement en protéines totales, aucun des additifs (en absence ou en présence de sorbitol) n'a d'effet probant. Pour cette souche, c'est l'addition de PEG 9% (sans sorbitol) ou de cuivre (avec sorbitol) qui induit une synthèse accrue de la forme mature de l'HSP22 recombinante (Figures 12A & 12B). Lamb et al. (2000) ont déterminé la structure tridimensionnelle (à une résolution de 2,75 Å) du grand domaine de la chaperone à cuivre de la superoxyde dismutase humaine. Ils ont mis en évidence la présence d'1 atome de zinc et de 0,2 atome de cuivre par molécule de chaperone. Rien ne prouve que la forme mature de l'HSP22 (recombinante ou dans la mitochondrie de feuille de pois) contient l'un et/ou l'autre de ces cations. Mais si tel est le cas, et à condition que la membrane bactérienne y soit perméable, l'addition de zinc ou de cuivre dans le milieu de culture pendant l'induction de l'expression peut s'avérer un élément de stabilisation de la structure de la protéine bio-synthétisée dans la bactérie. Une structure plus stable étant moins sujette à la dégradation par les protéases, la teneur en protéine recombinante est plus élevée dans ces conditions. |
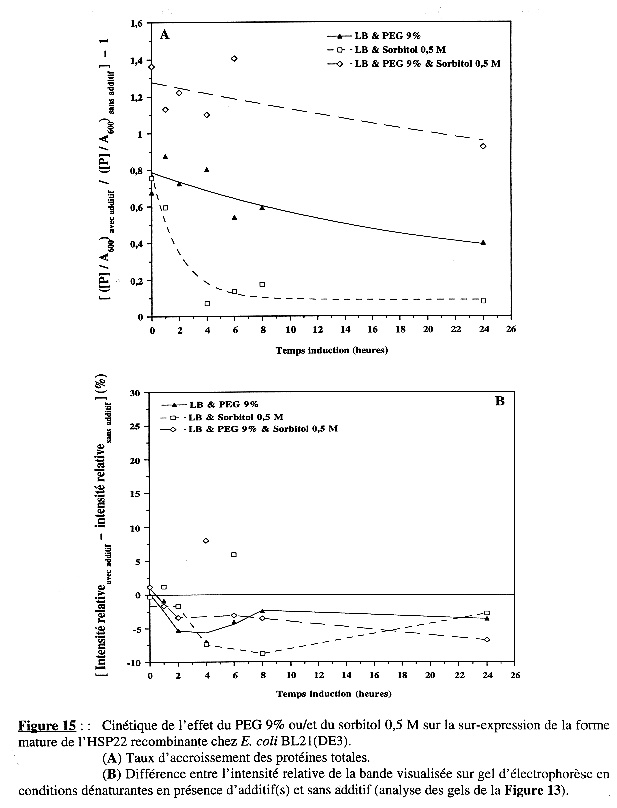
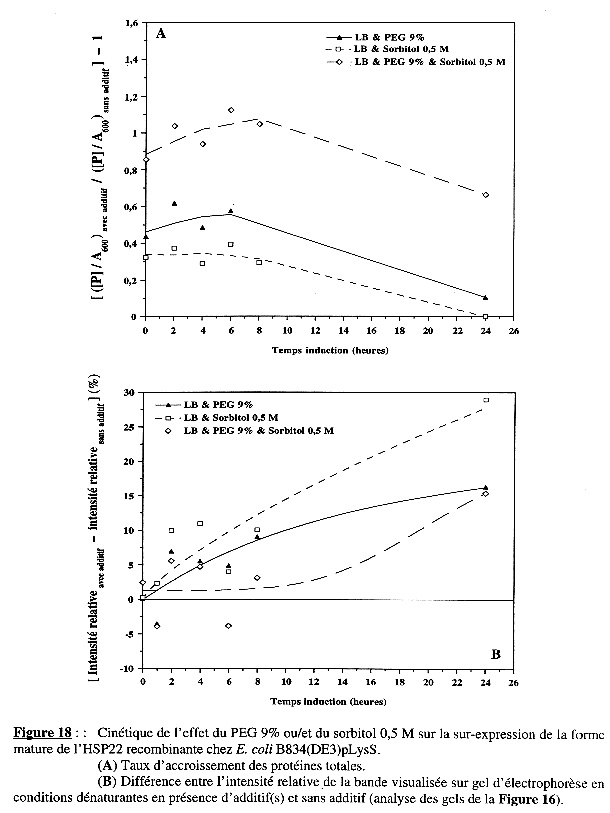
L'étude de la cinétique de l'effet du PEG 9% ou/et du sorbitol 0,5 M a montré, dans le cas de BL21(DE3), que le sorbitol diminue la quantité de protéines totales synthétisées et ralentit la vitesse de synthèse des protéines. Mais, la présence conjointe du PEG 9% abolit l'effet drastique du sorbitol et, inversement, le sorbitol potentialise celui du PEG. En revanche, l'un ou/et l'autre de ces additifs n'ont pas d'effet sur la synthèse spécifique de la forme mature de l'HSP22 recombinante (Figures 15B & 15B). Dans le cas de B834(DE3)pLysS, la cinétique du taux d'accroissement des protéines totales dans les trois conditions étudiées est la même que celle observée avec BL21(DE3). Mais, le résultat le plus intéressant en ce qui concerne l'effet des additifs a trait à la synthèse spécifique de la protéine sur-exprimée chez B834(DE3)pLysS : le PEG 9% et surtout le sorbitol 0,5 M induisent une synthèse apparente élevée de la forme mature de l'HSP22 recombinante, au moins pendant les 6 premières heures d'induction de l'expression (Figures 18A & 18B). |
2. Purification partielle de l'HSP22 recombinante De manière générale, les conditions choisies ne sont pas encore optimales. La forme mature de l'HSP22 recombinante se retrouve en trop grande proportion dans les surnageants obtenus à chaque étape (exception faite du surnageant après la dialyse). Il en résulte un rendement trop faible. En ce qui concerne l'état de pureté de la protéine à chaque étape, nous n'avons que le seul critère de l'intensité relative pour la bande correspondant à cette protéine. L'obtention d'un anticorps dans l'optique de Western-Blot est donc une priorité. Malgré tout, la quantité de protéine soluble (contenue dans le surnageant après la dialyse), donc probablement repliée correctement, est non négligeable (surtout pour BL21(DE3)). Améliorant les étapes qui précèdent, il est probable que cette quantité augmente pour les deux souches. C'est d'autant plus important que les étapes de purification que nous envisageons à la suite de celles décrites dans ce travail sont une chromatographie d'échange d'ions suivie d'une chromatographie sur gel de filtration. Dans les deux cas, il est préférable (du point de vue de la manipulation et du rendement) d'avoir à traiter un échantillon qui contient une protéine soluble. En d'autres termes, qu'il ne soit pas nécessaire de re-dénaturer la protéine pour effectuer ces étapes de purification. |
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}