
| Respiration mitochondriale - Etude de l'action d'inhibiteur et d'agent découplant |
| Tweet |
|
| Voir un développement sur l'oxygraphe à électrode à oxygène - l'intensité et le quotient respiratoire |
|
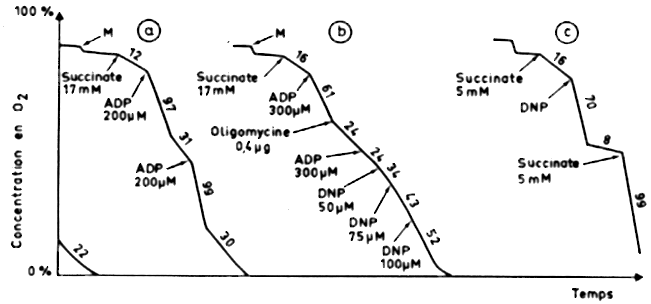
Analyser les oxygrammes (figure ci-dessous) et en déduire le mode d'action de l'oligomycine et du dinitrophénol (DNP).
|
|
L'intensité respiratoire (IR) est la quantité d'oxygène consommé (ou de CO2 dégagé) par unité de temps et de matière biologique (masse, nombre, surface, ...). Exemple : IR = 12 pmoles O2 / min / mg protéine La dépendance de la vitesse de respiration en fonction de la disponibilité de l'ADP (substrat de l'ATP synthase) est appelé contrôle respiratoire. IR
en présence d'ADP |
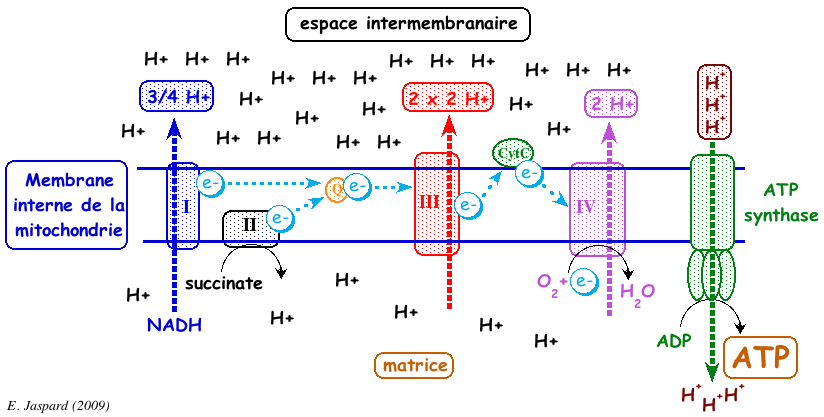
Ci-dessous : schéma général d'un fonctionnement correct de la chaîne respiratoire :
|
| Additif | Gradient de protons | Transfert d'électrons | Consommation d'O2 | Remarque |
| ADP absent | fort | ralenti | ralentie | Les protons sont plus difficilement expulsés par les complexes I, II et IV. |
| ADP présent | faible : formation d'ATP (les protons retournent dans la matrice mitochondriale) | accéléré | augmente | Le mécanisme d'action de l'ATP synthase dissipe le gradient de protons : l'ADP est phosphorylé en ATP à raison de 3 protons par molécule d'ATP synthétisée. |
| Agent découplant | annulation : il n'y a plus synthèse d'ATP | maximal | maximale | ----- |
|
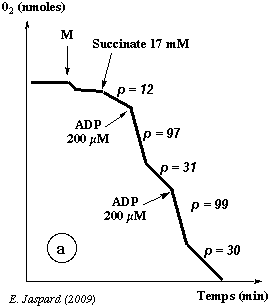
Enregistrement a M = les mitochondries purifiées sont ajoutées au milieu : on observe une pente non négligeable de courte durée. Cette consommation transitoire d'oxygène en absence de substrats de la chaîne respiratoire (le succinate n'est pas encore ajouté) est liée aux substrats endogènes contenus dans les mitochondries elles-mêmes. Leur quantité étant trés faible, la consommation d'oxygène ne dure pas.
|
| Additif | Effet |
| Succinate 17 mM : p = 12 | Les électrons portés par le succinate entrent dans la chaîne respiratoire au niveau du complexe II. Ces électrons sont transférés jusqu'au complexe IV où l'oxygène est réduit en eau : O2 + 4 H+ + 4 e- -> 2 H2O |
| ADP 200 µM : p = 97 | L'ADP est le substrat de l'ATP synthase. La chaîne respiratoire fonctionne davantage (donc l'oxygène est davantage consommé) pour reconstituer le gradient de protons (force proton-motrice) qui est dissipé par l'ATP synthase quand elle synthétise l'ATP. |
| p = 31 | Tout l'ADP ajouté a été transformé en ATP. L'ATP synthase ne fonctionne plus, donc il n'y a plus dissipation du gradient de protons. L'expulsion de protons de la matrice vers l'espace intermembranaire est plus difficile, donc la chaîne respiratoire ralentit. En conséquence, la consommation d'oxygène ralentit. |
| ADP 200 µM : p = 99 | Même effet qu'après la 1ère addition d'ADP 200 µM (p = 97). |
| p = 30 | Tout l'ADP ajouté a été transformé en ATP. |
| Conclusion : l'enregistrement a démontre le couplage respiratoire dans les mitochondries. | |
|
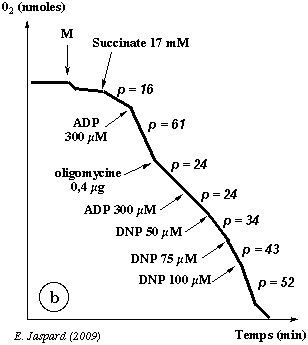
Enregistrement b
|
| Additif | Effet |
| Succinate 17 mM : p = 16 | Voir enregistrement a. |
| ADP 300 µM : p = 61 | Voir enregistrement a. |
| Oligomycine 0,4 µg : p = 24 | L'addition d'oligomycine inhibe l'accélération transitoire de la consommation d'oxygène
due au couplage respiratoire après addition de l'ADP (la pente
= 61 passe à 24). Il y a alors un excès d'ADP "inactif", c'est-à-dire qui ne peut être transformé en ATP. |
| ADP 300 µM : p = 24 | Même effet qu'après la 1ère
addition d'ADP 200 µM (p = 97). |
| DNP 50 µM : p = 34 |
L'addition de concentrations croissantes de DNP restaure progressivement la consommation d'oxygène. Attention : il n'y pas restauration du couplage respiratoire (comme on le voit après). |
On peut formuler 2 hypothèses :
|
|
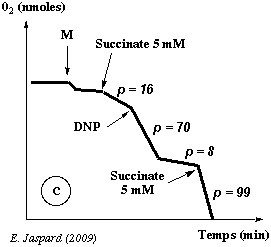
Enregistrement c
|
| Additif | Effet |
| Succinate 5 mM : p = 16 | Voir l'enregistrement a. |
| DNP : p = 70 | Sans addition d'ADP (donc en condition limitante en ADP où seul l'ADP endogène des mitochondries est disponible), il y a malgré tout une augmentation de la consommation d'oxygène en présence de DNP. |
| p = 8 | Cette consommation ne s'arrête que quand tout le substrat (ici le succinate) a été consommé. |
| Succinate 5 mM : p = 99 | -------- |
Conclusion : l'enregistrement c permet donc de retenir l'hypothèse d'un découplage de la chaîne respiratoire et de la synthèse d'ATP. |
|
|
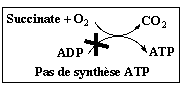
Oligomycine : inhibition de la synthèse d'ATP
|
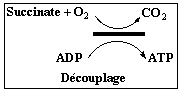
Dinitrophénol ou DNP : découplage
|
|
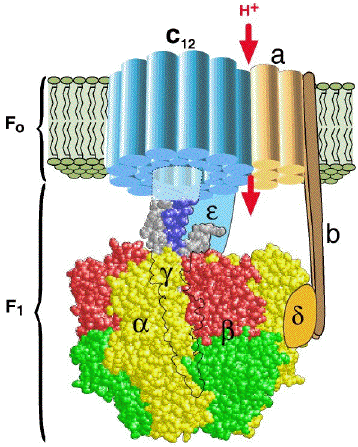
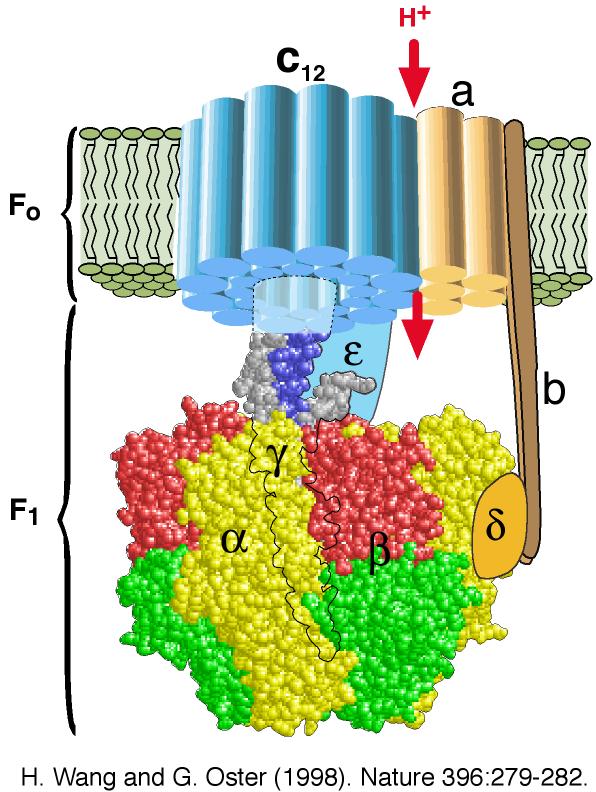
Complément sur les inhibiteurs de la phosphorylation (exemple : oligomycine). Ils empêchent la consommation d'oxygène après l'addition d'ADP mais n'ont pas d'effet sur la stimulation de la respiration par les agents découplants. Le complexe V ou ATP synthase F0F1, se sert du gradient de concentration de protons comme source d'énergie pour synthétiser l'ATP :
Source : Wang, H, & Oster, G. (1998) Nature 396, 279 - 282
|
|
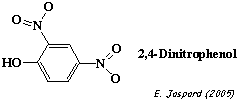
Complément sur les agents découplants (exemple : 2,4 dinitrophenol).
Ils dissipent artificiellement le gradient de protons et il n'y a plus synthèse d'ATP. Le transfert d'électrons et la consommation d'O2 sont à leur maximum. Le carbonyl cyanide m-chloro-phenyl hydrazone (CCCP) est un composé lipidique faiblement acide (figure ci-contre). C'est un agent découplant trés fort. Dans la forme ionisée du CCCP, la charge négative est délocalisée sur environ 10 atomes. Le champs électrique qui entoure la forme anionique est donc trés faible. Celà permet à l'anion de diffuser librement dans un milieu non polaire tel que la membrane (bicouche phospholipidique). Le p-trifluoromethoxy-carbonyl-cyanide-phenyl hydrazone (FCCP) est similaire au CCCP.Pour plus de détails, aller au site : "Uncouplers and Inhibitors" |
![]()
{kind=link}