| EST de cerveau d'abeille - Analyse de l'article de Whitfield et al. (2002) |
| Tweet |
|
|
1. Introduction 2. Exemple de protocole pour la préparation d'une banque normalisée et soustraite en une seule étape 3. L'épissage alternatif et ses conséquences sur les EST obtenues 4. Analyse des EST obtenues à partir du cerveau de l'abeille |
5. Logiciels et bases de données utilisés 6. Puces à ADN 7. Résumé des résultats 8. Liens Internet et références bibliographiques |
|
1. Introduction L'article a été publié en 2002. Or le génome de l'abeille n'a été publié qu'en 2006. C'est la raison pour laquelle, dans cet article, l'annotation des gènes de l'abeille a été faite par rapport au génome de la Drosophile.
Source : Genome.gov
Le génome de l'abeille (environ 200 méga-bases) a été séquencé par une méthode en vrac ("shotgun") et clonage dans des BAC. Il est constitué de 16 chromosomes et 1 chromosome mitochondrial. Les populations d'abeilles déclinent dans le monde entier. Par exemple, les apiculteurs américains perdent 30% de leurs colonies annuellement. Les abeilles sont des pollinisateurs critiques de nos cultures : ces pertes ont des conséquences majeures pour la viabilité de notre industrie agricole. |
Vie et comportement social de l'abeille Les abeilles sont des insectes sociaux, qui vivent en colonies qui peuvent contenir des dizaines de milliers d'individus. La vie de la colonie est organisée via une sub-division des tâches complexe et sophistiquée.
Ces tâches sont réparties parmi les individus via le processus de maturation du comportement ("behavioral maturation ") :
Intéret d'étudier l'abeille : étude de la "plasticité" neuronale et comportementale de l'abeille en relation avec son comportement social, ses capacités d'apprentissage et de mémorisation. Perspectives : implications des résultats en écologie et dans le domaine de l'évolution des espèces. Par extension, on peut mentionner l'importance économique de la pollinisation des fleurs et de la fabrique du miel par ce type d'abeille. Justification de l'obtention d'un trés grand nombre d'EST ("expressed sequence tag") :
Matériel de départ : 600 cerveaux d'abeilles ouvrières d'âges différents (de 1 à 30 jours) ont été isolés pour l'obtention des ARN à l'origine des banques.
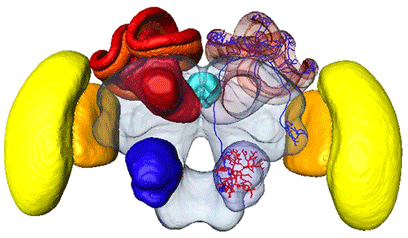
Source : Virtual Atlas of the Honeybee Brain |
|
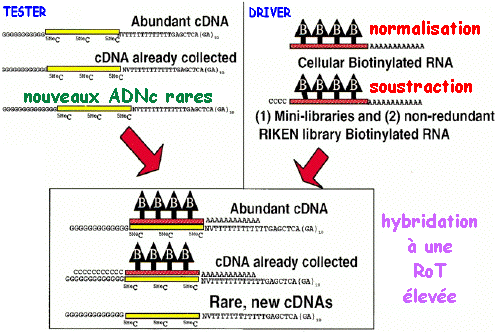
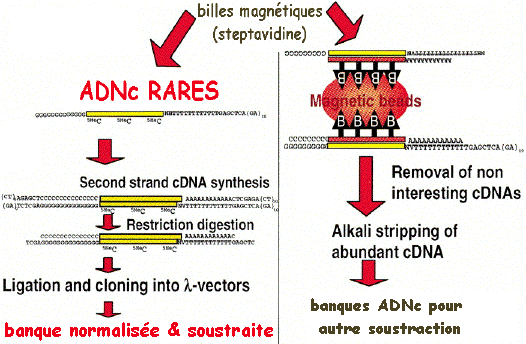
2. Exemple de protocole pour la préparation d'une banque normalisée et soustraite en une seule étape (Carninci et al., 2000) Technique mise au point à l'institut "RIKEN" (Japon). La stratégie repose sur :
La normalisation et la soustraction sont effectuées en une seule étape.
L'hybridation entre acides nucléiques dépend de nombreux paramètres physico-chimiques. En particulier, la probabilité d'hybridation entre des séquences complémentaires augmente avec le temps de la réaction et la concentration en acides nucléiques. Pour tenir compte de ces 2 facteurs, on définit le produit (concentration x temps) appelé :
La valeur RoT 1/2 correspond à 50% de molécules hybridées. Banque d'ADNc de tailles sélectionnées ("size-selected cDNA library").
Les ADNc sont séparés sur gel d'agarose. Puis la partie du gel correspondant aux ADNc de tailles désirées est découpée et les ADNc sont élués du gel. |
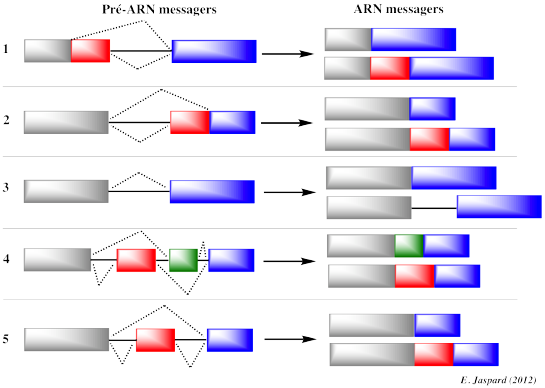
3. L'épissage alternatif et ses conséquences sur les EST obtenues Les génes sont transcrits sous forme d'ARN messagers pré-matures (synonymes : transcrits primaires ou pré-ARNm) qui contiennent des introns (séquences de l'ARN pré-mature non retenues dans la séquence finale de l'ARN messager qui code la protéine) et des exons qui sont assemblés selon différentes combinaisons (épissage alternatif). Les introns ont des tailles extrêmement variables : de plusieurs centaines de nucléotides à plusieurs centaines de milliers de nucléotides. L'épissage alternatif est le processus qui permet à un même gène de générer différents transcrits selon la combinaison des exons qui formeront l'ARN messager mature. L'épissage est effectué par deux réactions de trans-estérification au sein de complexes appelés spliceosomes formés, entre autres, de 5 particules ribonucléoprotéiques appelées SnRNP ("Small nuclear RiboNucleoProtein"). Ce sont des protéines associées à des petits ARN nucléaires ("small nuclear RNA" - snRNA) riches en uracile (U1, U2, U4, U5 et U6). Figure ci-dessous : Les 5 types d'épissage alternatif
|
Les répercutions de l'épissage alternatif sur les EST que l'on peut obtenir sont les suivantes :
Figure adaptée de "Précis de génomique", Gibson & Muse (2004)
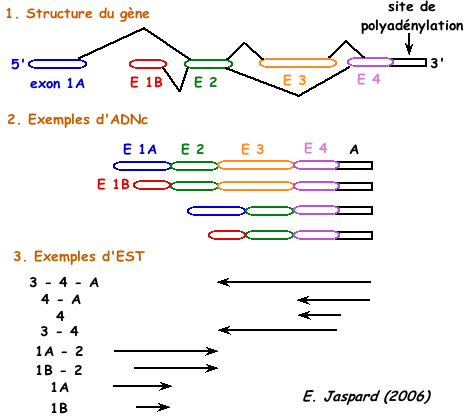
Puisque les séquences des EST ne correspondent qu'aux extrémités 3' ou 5' des ADNc, des séquences distinctes d'EST issues d'un même gène peuvent être interprétées dans un premier temps comme issues de gènes différents. La comparaison des EST et des séquences d'ADN génomique permet de lever cette ambiguïté et d'associer différents EST à un gène unique. |
| Voir un cours sur le spliceosome. |
|
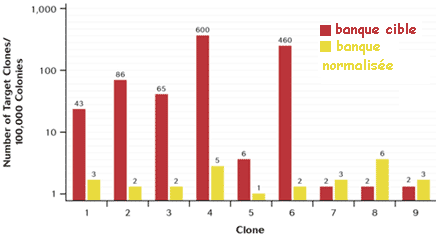
4. Analyse des EST obtenues à partir du cerveau de l'abeille Remarque 1 : dans cette étude, la banque a été normalisée puis soustraite. Remarque 2 : en 2012, la base de données UniGene (NCBI) contient plus de 190.000 EST et ARNm d'abeille regroupées en plus de 24.000 groupes ou "clusters". |
| Nombre total de séquences EST | 21408 |
| Nombre total de clones ADNc séquencés (5'EST) | 20256 |
| 5'EST issues de la banque normalisée | 7968 |
| 5'EST issues de la banque normalisée puis traitée par hybridation soustractive | 12288 |
| Séquences 5'EST redondante | 960 |
| Séquences 3'EST | 192 |
| Nombre total de séquences d'EST
de haute qualité taille moyenne : 494 paires de base |
15311 |
| Nombre d'EST initiales pour l'assemblage | 9481 |
| Nombre de contig | 3136 |
| Nombre de singletons | 5830 |
| Nombre de transcrits supposés
après regroupement des EST (Résultats conservés issus de CAP3) |
8966 - 54 = 8912 |
Qualité des EST et correspondance avec des gènes
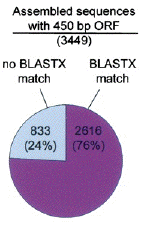
Source : Whitfield et al. (2002) a. L'analyse avec BLAST (e-value ≤ e-5) indique que, parmi ces 8912 séquences assemblées, 3501 (= 2616 + 885 / 39%) ont un homologue connu dans la base de données "Non-Redundant Protein (nr)". b. Sur les 8912 séquences assemblées, 3449 possèdent une phase de lecture ouverte ("Open Reading Frame - ORF") d'au moins 450 paires de bases. Parmi ces séquences, 2616 (76%) ont un homologue connu dans la base de données "nr". En d'autres termes, 833 (24%) gènes possibles codant des protéines et exprimés dans le cerveau de l'abeille n'ont pas d'homologues dans la base de données "nr" et seraient donc nouveaux.
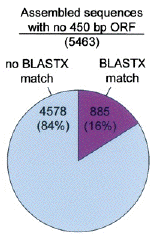
Source : Whitfield et al. (2002) c. Sur les 8912 séquences assemblées, 5463 possèdent une ORF inférieure à 450 paires de bases. Parmi ces séquences, 885 (16%) ont un homologue connu dans la base de données "nr". d. En ce qui concerne les 4578 (84%) autres séquences qui ne correspondent à rien, diverses explications sont fournies :
|
|
Comparaison avec d'autres espèces a. L'abeille et la drosophile sont toutes deux des arthropodes. En effet, l'analyse avec BLAST indique que, parmi les 3501 séquences assemblées qui ont un homologue connu dans la base de données "nr", 2245 (64%) sont similaires de séquences de protéines de la drosophile. b. Une comparaison avec BLASTX indique que 3362 séquences d'EST assemblées de l'abeille sont homologues à 2672 séquences de la drosophile soit (19,6% de redondance). Sur la base d'une redondance d'environ 20% dans les 8912 séquences assemblées, les auteurs estiment que ces 8912 séquences représenteraient 7100 gènes exprimés. Ils en concluent que : si l'abeille a le même nombre de gènes que la drosophile, ces 7100 gènes représenteraient 50% des gènes du génome de l'abeille. Rappel : l'article a été publié en 2002. Or le génome de l'abeille n'a été publié qu'en octobre 2006. Annotation fonctionnelle des séquences d'EST assemblées du cerveau d'abeille Elle a été faite :
Résultats :
*En 2001, seuls 42 gènes de la drosophile étaient annotés comme liés au comportement. Pour augmenter ce nombre, les auteurs ont généré une liste de 106 gènes à partir de mutants de la drosophile ayant des modifications de différents aspects du comportement. |
| Aller à la base de donnée : Honey Bee Genome Project |
5. Logiciels et bases de données utilisés
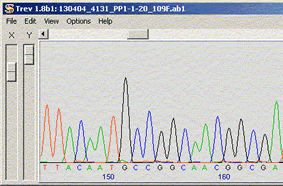
Electrophoregramme obtenu par appel de base
Analyse du bruit de fond par le programme GenePix |
|
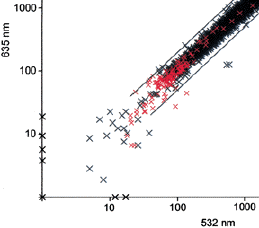
6. Caractéristiques des échantillons pour les puces à ADN Le but de cette analyse n'est pas une analyse transcriptomique classique, c'est-à-dire une comparaison des gènes différentiellement transcrits entre une condition témoin et une condition pathologique. Cette analyse a pour but de valider le point suivant : la quantité d'ARN extraite d'UN cerveau d'abeille est-elle suffisante pour toute expérience ultérieure d'hybridation aux 2 longueurs d'onde (635 nm et 532 nm) ? En effet, les ARN sont extraits de 2 cerveaux d'abeille puis l'échantillon est divisé en 2 aliquotes. Chacun est marqué par un fluorophore (Cy3 et Cy5). Les échantillons marqués sont mélangés et hybridés à la même puce. La fluorescence des puces sont lues aux 2 longueurs d'onde. Les résultats du tableau 7 et de la figure 2 confirment la possibilité de ne travailler que sur un cerveau. Choix des EST
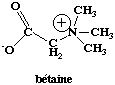
Bétaine
La relation entre température de fusion et la composition en base des sondes est moins stricte en présence de bétaine. Ainsi, l'emploi de la bétaine dans les puces à ADN augmente (Diehl et al., 2001) :
|
|
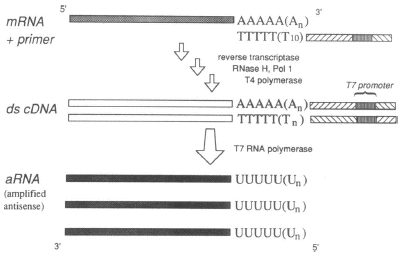
Cibles Les cibles sont des ARNm extraits d'un mélange de 2 cerveaux d'abeilles adultes puis amplifiés. Le marquage est effectué au cours de la rétro-transcription des ARN : un échantillon aliquote est marqué au Cy3 et un autre échantillon aliquote est marqué au Cy5. Les 2 échantillons sont ensuite mélangés et hybridés aux sondes (p 560). Amplification d'ARN : cette technique est utilisée pour la synthèse et le marquage de cibles pour des expériences de puces, quand la quantité de matériel de départ est limitante. Un oligonucléotide qui contient la séquence du promoteur de l'ARN polymérase T7 est incorporé au cours de la synthèse du second brin d'ADNc. L'ARN polymérase T7 permet ensuite la synthèse d'un ARN antisens amplifié ("amplified antisense RNA" - "aRNA"). Remarque : dans cette étude, l'émission de fluorescence est suivie à 532 nm et 635 nm.
Source : Van Gelder et al., 1990 Contrôles
Données issues des puces à ADN
Les valeurs d'intensité de fluorescence ont été normalisées de sorte que la moyenne du rapport des intensités (635 / 532 nm) = 1. Les résultats des puces à ADN (tableau ci-dessous) confirment que la grande majorité des EST du cerveau sont issues de transcrits réellement exprimés dans le cerveau de l'abeille. |
| Longueur d'onde | 635 nm | 532 nm |
| Nombre total de spots abeille | 7329 | 7329 |
| spots abeille > spots contrôles + 2 fois déviation standard | 6647 (91%) | 6631 (90%) |
|
Les transcrits potentiels ont été annotés sur la base des données disponibles en 2002 pour la drosophile.
Conclusions - perspectives Beaucoup de phénomènes liés au comportement chez l'abeille ne sont pas observés chez la drosophile. De plus l'abeille est haplo-diploïde et a le taux de recombinaison le plus élevé connu chez les animaux. Séquence abeille - gène du comportement : CAM Kinase II / Fichier Unigene BB170005A10D08 Pour mieux corréler l'expression de certains gènes au comportement de l'abeille :
Exemples de continuité de ce travail : Nunes et al. (2004); Whitfield et al. (2006). Approches complémentaires plus récentes :
|
| 8. Liens Internet et références bibliographiques |
|
Ressources génomiques et transcriptomiques Honey Bee Genome Project & Apis mellifera (honey bee) genome view "The Virtual Atlas of the Honeybee Brain" BeeBase "Apis mellifera transcriptome sequencing project" |
|
|
Carninci et al. (2000) "Normalization and subtraction of cap-trapper-selected cDNAs to prepare full-length cDNA libraries for rapid discovery of new genes" Genome Res. 10, 1431 - 1432 Whitfield et al. (2002) "Annotated Expressed Sequence Tags and cDNA Microarrays for Studies of Brain and Behavior in the Honey Bee" Genome Res. 12, 555 - 566 Van Gelder et al. (1990) "Amplified RNA synthesized from limited quantities of heterogeneous cDNA" Proc. Natl. Acad. Sci. U S A. 87, 1663 - 1667 Diehl et al. (2001) "Manufacturing DNA microarrays of high spot homogeneity and reduced background signal" Nuc. Acids Res. 29, e38 |
|
|
Nunes et al. (2004) "The use of Open Reading frame ESTs (ORESTES) for analysis of the honey bee transcriptome" BMC Genomics 5, 84 Whitfield et al. (2006) "Genomic dissection of behavioral maturation in the honey bee" Proc. Natl. Acad. Sci. USA 103, 16068 - 16075 Ament et al. (2010) "Nutritional regulation of division of labor in honey bees: toward a systems biology perspective" Systems Biology and Medicine 2, 566 - 576 Greenberg et al. (2012) "Behavioral plasticity in honey bees is associated with differences in brain microRNA transcriptome" Genes, Brain and Behavior 11, 660 - 670 Baker et al. (2012) "Age-related learning deficits can be reversible in honeybees Apis mellifera" Exp. Gerontology 47, 764 - 772 |
![]()