Purification de la glutamate deshydrogénase EC 1.4.1.4 de la semence de Pisum sativum - Marie-Hélène Guilleux - 2004 Méthodes |
|
Voir un ensemble de cours de biochimie. Voir un ensemble d'exercices pour travaux pratiques de Biochimie (item 21). |
|
1. Mesure de l'activité aminante et désaminante de la GDH et dosage des protéines totales L'activité aminante de la glutamate deshydrogénase dépendante du NADPH est mesurée en suivant la consommation du NADPH par spectrophotométrie à 340 nm, à 30°C. Le mélange réactionnel contient : |
| HEPES | 50 mM, pH6.8 |
| α-cétoglutarate | 20 mM |
| NH4Cl | 50 mM |
| NADPH | 0.2 mM |
| Mélange enzymatique | X mL |
| Eau distillée | jusqu'à 1mL |
Mesure de l'activité désaminante : pour savoir si l'activité aminante mesurée n'est pas celle de l'isoforme [EC 1.4.1.2] qu'elle peut catalyser de façon non préférentielle, l'activité désaminante est également mesurée sur chaque échantillon. C'est la formation du NADH qui est alors suivie à 340 nm et à 30°C. Pour cette réaction, le substrat est le glutamate, le coenzyme est NAD 50 mM dans le tampon Hepes 50 mM pH 6.8. La concentration en protéines totales est déterminée selon la technique de Bradford (1976) en utilisant l'albumine de sérum bovin comme standard. |
2. Purification de la GDH EC 1.4.1.4 a. Matériel biologique - Extrait brut Le matériel biologique utilisé est des semences de pois (Pisum sativum) imbibées pendant 12 et 22 heures. L'extrait brut est obtenu en broyant des semences de pois selon un rapport 1g de semences dans 4 mL de tampon d'extraction. Le tampon d'extraction contenant 50 mM Hepes pH7.5, 2 mM EDTA, 1 mM β-mercaptoéthanol, 1 mM PMSF. L'extrait brut est clarifié par centrifugation (30 min, 27 000g, 4°C). On récupère le surnageant. b. Précipitation au sulfate d'ammonium Le surnageant est précipité par du sulfate d'ammonium (70% saturation), puis centrifugé (30 min, 27000g, 4°C). Le culot récupéré est repris dans un volume minimal de tampon d'extraction avec 20% de saturation de sulfate d'ammonium et dialysé contre 5L de tampon d'extraction, toute une nuit à 4°C. |
c. Chromatographie échangeuse d'anions DEAE-cellulose Le dialysat est alors chargé sur un gel de chromatographie échangeuse d'anions DEAE-cellulose, équilibré avec du tampon d'extraction et élué par un gradient de NaCl (0 à 0.5 M) dans du tampon d'extraction. Des fractions de 1 mL sont collectées avec un débit de 0.3 mL/min. Les fractions actives sont ensuite réunies. d. Chromatographie d'affinité pigments L'éluat actif est chargé sur un gel de chromatographie d'affinité pigments, équilibré avec du tampon d'extraction et élué par un gradient de NaCl (0 à 1 M) dans du tampon d'extraction. Des fractions de 1 mL sont collectées avec un débit de 0.24mL/min. Les fractions actives sont réunies. e. Chromatographie d'affinité NAD/ATP L'éluat actif est chargé sur un gel de chromatographie d'affinité NAD/ATP équilibré avec du tampon d'extraction (débit = 0.18 mL/min). L'activité est récupérée dans la charge et le rinçage. La charge et le rinçage sont réunis et concentrés contre 2L de PEG (polyéthylène glycol). Ceci permet de récupérer l'activité dans un petit volume. |
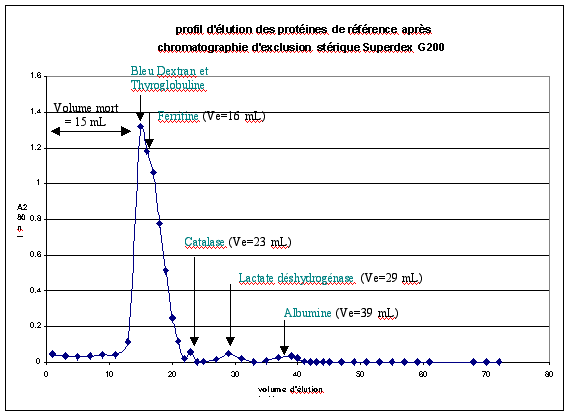
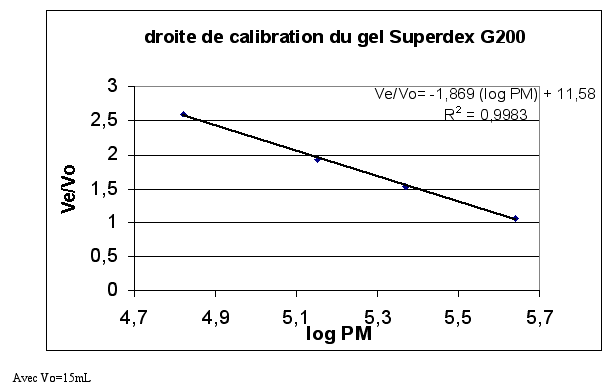
f. Chromatographie d'exclusion stérique Superdex G200 Ce type de chromatographie permet non seulement de séparer les protéines mais également de déterminer le poids moléculaire des protéines. Afin de déterminer la masse molaire (MM) des protéines éluées, des protéines de référence encore appelées marqueurs de MM connus sont déposées sur le gel. Il s'agit de la thyroglobuline (669 kDa), la ferritine (440 kDa), la catalase (232 kDa), la lactate déshydrogénase (140 kDa) et l'albumine (66 kDa). Le domaine de fractionnement du gel Superdex G200 est compris entre 104 et 6.105 Da. Ainsi, en utilisant le Bleu Dextran, de très haut poids moléculaire, 2.106 Da, le volume mort Vo peut être déterminé. Ici, V0=15 mL. La thyroglobuline est éluée avec le Bleu Dextran car sa MM est supérieure au domaine de fractionnement. Le profil d'élutionest tracé après la mesure d'absorbance à 280 nm de chaque fraction éluée (fraction de 1 mL).
Le volume d'élution Ve de chaque protéine de référence est déterminé. La droite de calibration Ve/V0 = f (log MM) peut être tracée :
L'échantillon concentré au PEG est alors chargé sur le gel. L'élution se fait par passage du tampon d'extraction avec un débit de 0.14 mL/min. Puis la masse molaire des protéines éluées peut être déduite. |
g. Electrophorèse SDS-PAGE Le principe de l'électrophorèse SDS-PAGE est de traiter l'échantillon protéique avec un agent réducteur (β-mercaptoéthanol) pour rompre les ponts disulfures et un détergent anionique (SDS) qui dénature les protéines et leur confère une enveloppe de charge négative (Hames B.D. et al., 2000). L'échantillon est fractionné par une électrophorèse sur gel de polyacrylamide 10%. Tous les fragments protéiques sont séparés selon leur masse molaire qui est déterminé grâce à des marqueurs. Les protéines standards utilisées sont des marqueurs de bas poids moléculaires contenant la phosphorylase b (97 kDa), l'albumine (66 kDa), l'ovalbumine (45 kDa), l'anhydrase carbonique (30 kDa), l'inhibiteur de trypsine (20,1 kDa), et l'α-lactalbumine (14,4 kDa). Cette technique permet de déterminer la masse molaire des sous-unités formant une protéine, et de se rendre compte du degré de purification atteint à chaque étape. |
![]()