| Étude du facteur de croissance du nerf ("Nerve growth factor" - NGF) |
| Tweet |
|
|
1. Énoncé 2. Préambule pour la compréhension de l'exercice 3. Réponse partie A 4. Réponse partie B |
5. Synthèse des résultats et complément d'informations
6. Liens Internet et références bibliographiques |
|
1. Énoncé Le facteur de croissance du nerf béta ou "β-nerve growth factor" ou β-NGF est une hormone de croissance qui contrôle la différenciation et le maintien de certains neurones. Chez la souris, cette protéine est produite par la glande sous-maxillaire. Le β-NGF contient un grand nombre d'acides aminés soufrés, on peut donc étudier sa biosynthèse in vivo en mesurant l'incorporation de 35S-cystéine dans des coupes de glandes. Partie A Après incubation de coupes de glandes en présence de 35S-cystéine, on extrait les protéines solubles. Elles sont déposées sur un gel d'immuno-adsorbant constitué par des anticoprs anti-NGF immobilisés sur des billes d'agarose. Les produits retenus par l'immuno-adsorbant sont élués et analysés par électrophorèse sur gel de polyacrylamide en conditions dénaturante (SDS-PAGE). Cette expérience (extraction, fixation/élution, électrophorèse) est réalisée après incubation des coupes avec de la 35S-cystéine pendant 1 heure puis addition d'un excès de cystéine froide (non radioactive) pendant un temps variable (technique de poussée-chasse ou "pulse-chase").
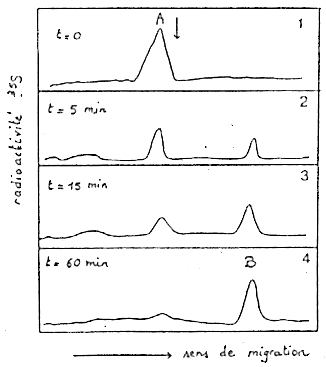
Les profils d'électrophorèse (figure 1 ci-dessous) montrent l'évolution de la radioactivité à divers temps suivant la chasse par la cystéine froide. La flèche indique la position de la chaîne catalytique de l'aspartate transcarbamylase (MM = 34000 Da).
Partie B 1. Le β-NGF est obtenu à l'état pur.
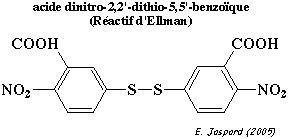
2. Le β-NGF ne réagit avec le réactif d'Ellman (dinitro-2,2'-dithio-5,5'-benzoïque - DNTB) qu'après traitement au β-mercaptoéthanol :
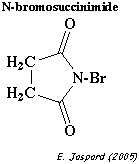
3. La réaction du β-NGF avec le N-bromo-succinimide (NBS) se traduit par une baisse de A280. La disparition d'1 mole du résidu d'acide aminé qui réagit avec le NBS correspond à ΔεM = 4000 M-1.cm-1. Ainsi le β-NGF à 1,25 mg/mL traité par le NBS done une absorbance de 2 au début de la réaction et de 0,8 à la fin de la réaction. 4. L'espèce A marquée par la 35S-cystéine (figure 1) est purifiée :
Sur la base de ces données, apporter toutes les précisions sur la structure primaire, tertiaire (liaison covalente) et quaternaire du β-NGF. |
|
2. Préambule pour la compréhension de l'exercice Le NGF ("high molecular weight 7S NGF complex") est constitué de 3 types de sous-unités => c'est le complexe [α-NGF]2 - [γ-NGF]2 - [β-NGF]2. |
| molécule | type | activité |
| sous-unité α-NGF | protéases à sérine de la famille des kallikréines | hydrolyse du précurseur β-NGF en β-NGF mature |
| sous-unité γ-NGF |
inhibe l'activité de la sous-unité α-NGF |
|
| précurseur β-NGF | --------- | |
| sous-unité β-NGF mature | homodimère (paire de feuillets β antiparallèles enroulés) avec 3 ponts disulfures et 2 atomes de zinc | médiateur des fonctions biologiques du NGF |
|
a. Marquage à la 35S-cystéine, technique de poussée-chasse Le souffre 35 est un émetteur β peu stable, principalement utilisé dans le marquage métabolique des protéines. En effet on peut obtenir facilement de la [35S]-méthionine ou cystéine avec des activités spécifiques extrêmement élevées. Cette méthode permet d'étudier la synthèse protéique in vivo par marquage avec des acides aminés marqués (voir un cours sur les radioéléments en biologie). Technique de poussée-chasse ("pulse-chase")
Cette technique a été utilisée notamment :
|
b. Immuno-adsorption et électrophorèse en conditions dénaturantes L'immuno-adsorbtion est une chromatographie d'affinité : des anticoprs anti-NGF immobilisés sur des billes d'agarose. Les produits retenus puis élués et analysés par SDS-PAGE (pics A et B de la figure 1) sont donc le NGF (ou des sous-unités du NGF). Les profils d'élution de la figure 1 mettent en évidence 2 espèces :
=> Hypothèse : la molécule B est une forme maturée de A. En d'autres termes, A est une forme précurseur de B. |
|
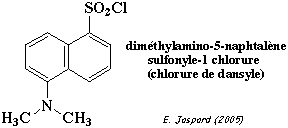
a. Masse molaire a. Les conditions dénaturantes dissocient les sous-unités d'un oligomère : conditions non dénaturantes => oligomère β-NGF = 24.000 Da; conditions dénaturantes => sous-unités β-NGF = 12.000 Da => Le β-NGF est un dimère. Réaction avec le chlorure de dansyle (DNS) :
Autres molécules pour marquer le résidu N-terminal d'une protéine : le chlorure de dansyle ou le chlorure de dabsyle (réactifs fluorescents), le fluoro-dinitrobenzène (réactif d'Edman) ou l'isocyanate de méthyle. Quantité de β-NGF et de DNS-Ser :
=> L'acide aminé en position N-terminale de chaque sous-unité β-NGF est une sérine. Réaction avec la carboxypeptidase Y : d'après l'alignement présenté ci-dessous, l'acide aminé à caractère basique en position C-terminale d'une sous-unité β-NGF serait une arginine. |
b. Le réactif d'Ellman (acide dinitro-2,2'-dithio-5,5'-benzoïque - DNTB) :
Il réagit avec les cystéines libres (groupements -SH). Les cystéines impliquées dans un pont disulfure doivent être préalablement réduites (traitement au β-mercaptoéthanol). Le chromophore formé est la DNTB-Cys.
a. La masse molaire du β-NGF est : MM = 24.000 Da = 24000 g/mol (voir un rappel sur le Dalton). b. [β-NGF] = 0,125 mg/mL = 0,125 g/L => [β-NGF] = (0,125 g/L / 24000 g/mol) = 5,2 10-6 M c. [DNTB-Cys] = (0,816 / 13600) = 60 10-6 M d. Nombre de DNTB-Cys = (60 10-6 / 5,2 10-6) = 12 Cys Conclusions
=> Il y a 3 ponts disulfure intra-chaînes ou 6 ponts disulfure inter-chaînes ou autre combinaison [intra + inter]. |
c. Réaction avec le N-bromo-succinimide (NBS)
Il coupe les protéines en réagissant avec les résidus tryptophane, tyrosine et histidine. A 280 nm, c'est la disparition du tryptophane que l'on suit.
a. La masse molaire du β-NGF est MM = 24.000 Da = 24000 g/mol. b. [β-NGF] = 1,25 mg/mL = 1,25 g/L => [β-NGF] = (1,25 g/L / 24000 g/mol) = 5,2 10-5 M c. Δ[Trp] = (0,8 / 4000) = 3 10-4 M d. Nombre de résidus tryptophane modifiés = (3 10-4 / 5,2 10-5) = 6 => Il y a 3 tryptophanes par sous-unité β-NGF. |
d. Réaction du chlorure de dansyle et action de la trypsine sur la molécule A L'acide aminé en position N-terminale de la molécule A est une sérine (comme dans le cas du β-NGF). La trypsine hydrolyse la liaison peptidique après les résidus basiques => juste avant la séquence en acide aminé de la molécule B, on doit trouver une arginine ou une lysine. |
|
5. Synthèse des résultats et complément d'informations a. Sous-unités α-NGF et γ-NGF (voir préambule)
L'alignement ci-dessous montre le trés fort taux d'identité entre les acides aminés des 2 types de sous-unités (encadrés en fond rouge).
Alignement généré avec MULTALIN - Figure générée avec ESPript |
b. Sous-unités précurseur β-NGF et β-NGF mature
Ci-dessous, alignement des 2 séquences en acides aminés :
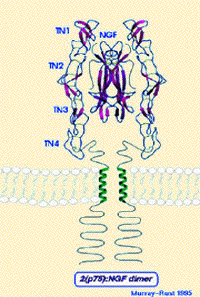
Figure ci-dessous : sous-unité β-NGF mature fixée à l'un de ses récepteurs
Source : "Le Cerveau (McGill)" |
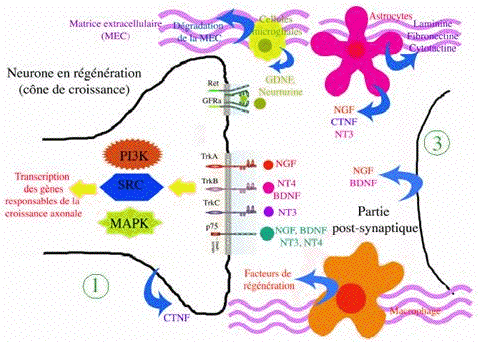
c. Aperçu de la réparation des lésions des neurones Des études chez les Vertébrés ont permis de caractériser des protéines qui interviennent à la suite d'une lésion du tissu nerveux (figure ci-dessous). Le facteur de croissance NGF et les facteurs neurotrophiques comme la neurotrophine 3 (NT3) et le GDNF ("glial cell line-derived neurotrophic factor") permettent la reconnexion d'axones endommagés chez le rat. Figure ci-dessous : schéma de synthèse des effecteurs impliqués dans la réparation nerveuse chez les Vertébrés.
Source : Laboratoire de Neuroimmunologie des Annélides De plus, deux protéines associées à la croissance axonale, GAP-43 ("growth associated protein") et CAP-43 ("cone associated protein"), permettent conjointement le bourgeonnement et l'élongation des axones. L'action de ces protéines est liée à la transcription accrue de leur gène respectif, induite par la lésion [Ramer et al. (2000), Bomze et al. (2001)]. |
| 6. Liens Internet et références bibliographiques |
|
Ramer et al. (2000) Nature 403, 312-316 Bomze et al. (2001) Nat Neurosci 4, 38-43 |
|
|
Facteurs de croissances non hématopoiétiques "Cell signaling" Gouet et al. (1999) "ESPript: multiple sequence alignments in PostScript" Bioinformatics 15, 305 - 308 |
|
|
Extrait de l'article : "Nerve growth factor: structure/function relationships" - Bradshaw et al. (1994) Protein Science 3, 1901 - 1913 Nerve growth factor which has a tertiary structure based on a cluster of 3 cystine disulfides and 2 very extended, but distorted beta-hairpins, is the prototype of a larger family of neurotrophins. Residues involved in these interactions and in the maintenance of tertiary and quaternary structure have been identified by chemical modification and site-directed mutagenesis, and this information can be related to their location in the 3-dimensional structure. For example, interactions between aromatic residues contribute to the stability of the NGF dimer, and specific surface lysine residues participate in receptor contacts. The conclusion from these studies is that receptor interactions involve broad surface regions, which may be composed of residues from both promoters in the dimer. |
|
Extrait de l'article : "Processing of the native nerve growth factor precursor to form biologically active nerve growth factor" Edwards et al. (1988) J Biol Chem 263, 6810 - 6815 In the mouse submaxillary gland β nerve growth factor (β-NGF) forms a complex with two members of the kallikrein family of serine proteases, termed the alpha and gamma subunits of NGF. We demonstrate that the β-NGF precursor produced in mammalian cells via a recombinant vaccinia virus can be cleaved by stoichiometric quantities of the γ-subunit to produce β-NGF. Trypsin in catalytic quantities also produces native β-NGF. Proper cleavage depends critically on the conformation of the precursor. β-NGF has at least 10-fold more biological activity than its precursor. |
|
Extrait de l'article : "Subunit interactions of the nerve and epidermal growth factor complexes: protection of the biological subunit from proteolytic modification" Nichols & Shooter (1985) Dev Neuroscience 7, 216 The nerve growth factor dimer (β-NGF) can undergo two proteolytic modifications, one near the amino terminus where a unique histidine/methionine bond is cleaved and the other at the carboxy terminus releasing the terminal arginine residue. The modification near the amino terminus, releasing the first eight amino acids as an octapeptide, occurs as the result of the action of a specific endopeptidase present in the submaxillary gland. The amino terminal octapeptide of beta NGF is protected by both the alpha and gamma subunits of 7S NGF and its loss has no effect on 7S NGF complex stabilization by zinc. The carboxy terminal arginine is protected only by the γ subunit. |
|
Synthèse de cette correction au format PDF (tous droits réservés biochimej). |
![]()