| Historique de l'évolution des modes d'alimentation en carbone et en énergie des êtres vivants |
| Tweet |
|
|
Source principale de ce cours : "Physiologie végétale" (1995) volume I - "Nutrition et métabolisme", Danielle Laval-Martin et Paul Mazliak, Collection "Méthodes", Hermann / ISBN : 2 7056 6253 7 |
|
Il existe 3 types de sources énergétiques utilisables par les êtres vivants :
Quel que soit le mode d'alimentation énergétique d'une cellule, l'énergie acquise y est stockée sous forme d'ATP, via une réaction de phosphorylation : ADP + Pi -> ATP Il existe 3 types de phosphorylation :
Il existe 3 catégories d'êtres vivants :
|
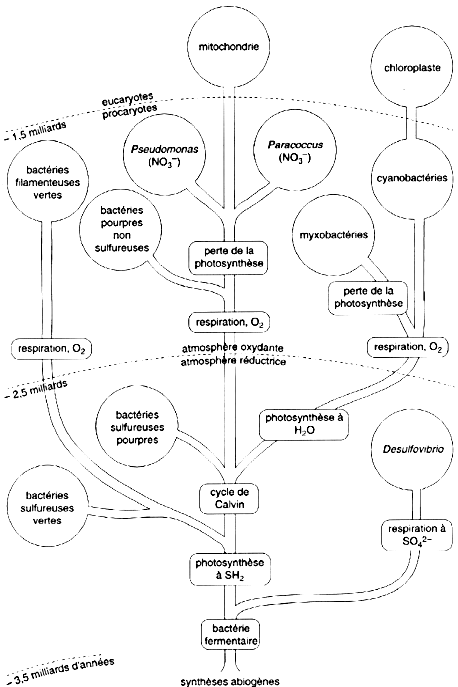
Ci-dessous : arbre phylogénique (sur la base des séquences du cytochrome c) retraçant l'évolution des métabolismes bactériens et notamment l'apparition de la respiration bien après la photosynthèse.
Source : "Physiologie végétale" (1995) Laval-Martin & Mazliak La respiration au sulfate a précédé la respiration à l'oxygène, apparue obligatoirement après la photosynthèse évoluée décomposant l'eau et enrichissant l'atmosphère en oxygène. La théorie endosymbiotique de l'origine de la cellule eucaryote postule que la mitochondrie dérive d'une bactérie respirante et le chloroplaste dérive des cyanobactéries.
Source : Futura-Sciences |
|
1. Fermentation (hétérotrophie primitive) Bactéries fermentaires (premiers êtres vivants). Le métabolisme carboné anaérobie associé aux fermentations se retrouve dans le cytoplasme des cellules eucaryotes. 2. Chimiosynthèse primitive (2 premiers milliards d'années de la terre) Bactéries aptes à oxyder plus complètement les aliments que les bactéries fermentaires.
|
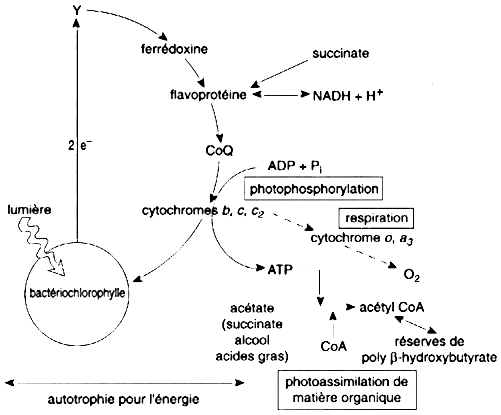
3. Photosynthèse bactérienne primitive Les pigments qui captent l'énergie lumineuse (bactériochlorophylle) ne forment qu'un seul photosystème (pigments et protéines). La source d'électrons est un composé minéral réduit (exemple : SH2). a. Photo - organotrophie de la bactérie pourpre non sulfureuse Rhodospirillum rubrum (figure ci-dessous) Photosystème à bactériochlorophylle qui opère une circulation cyclique des électrons délocalisés par les photons captés. Semi - autotrophie énergétique et dépendance quant aux matières organiques.
Source : "Physiologie végétale" (1995) Laval-Martin & Mazliak Deux évolutions importantes sont annoncées : l'ATP et le NAD+ réduit peuvent être utilisées pour réduire le CO2 de l'air. Donc, passage de la semi - autotrophie à l'autotrophie complète pour le carbone et l'énergie. Quand l'oxygène aura envahi l'atmosphère, les cytochromes c injecteront leurs électrons soit vers la bactériochlorophylle, soit vers d'autres cytochromes (a1, a3) et l'oxygène. La respiration apparaîtra comme "sous-produit" de la photosynthèse primitive. |
Réversion du cycle des acides tricarboxyliques Chez certains microorganismes anaérobies (par exemple, la delta-protéobactérie thermophile réductrice du soufre Hippea maritima), le cycle de l'acide tricarboxylique (TCA), y compris la réaction apparemment irréversible catalysée par la citrate synthase, peuvent être réversés et déboucher sur une fixation autotrophique du carbone.
Chez Hippea maritima, cette voie est induite par des pressions partielles élevées de CO2 : de telles pressions sont déterminantes pour transformer le produit acétyl coenzyme A en pyruvate (par carboxylation réductrice par la pyruvate synthase). Le cycle TCA oxydant reversé peut ainsi avoir fonctionné dans la fixation autotrophique du CO2 dans une atmosphère primordiale supposée riche en ce gaz. Voir Steffens et al. (2021) "High CO2 levels drive the TCA cycle backwards towards autotrophy" Nature 592, 784 - 788 |
b. Photosynthèse primitive des bactéries pourpres sulfureuses - (photo - autotrophie ou photolithotrophie primitives) Elles s'alimentent en énergie aux dépens du soleil en utilisant les photons captés par leur unique photosystème. Il y a photophosphorylation de l'ATP avec des composés soufrés comme source d'électrons. L'alimentation en carbone se fait par capture du CO2 atmosphérique réduit impliquant le cycle de Calvin. Le produit de la réduction du CO2 est un glucide de formule générale CH2O. La photosynthèse anaérobie rend ces bactéries autotrophes mais cette photosynthèse est primitive car la source d'électrons (exemple : SH2) n'est pas répandue à la surface de la terre. L'équation générale de la photosynthèse primitive est la suivante : lumière |
4. Photosynthèse évoluée des cyanobactéries, algues vertes, plantes vasculaires
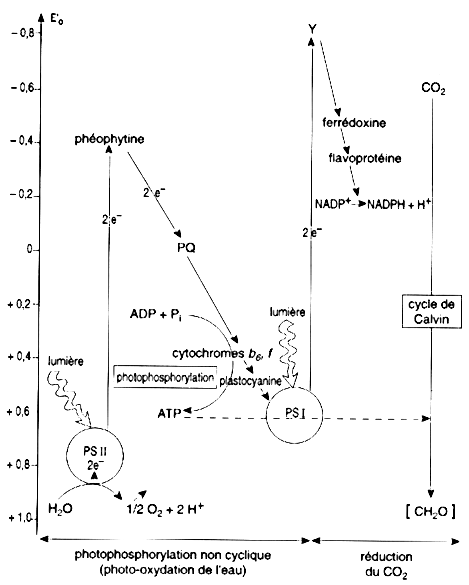
Source : "Physiologie végétale" (1995) Laval-Martin & Mazliak Quand les cyanobactéries (ou "algues bleues") ont remplacé les donneurs d'électrons soufrés par l'eau, les êtres photosynthétiques ont pu dés lors conquérir tous les milieux. On note :
L'équation générale de cette photosynthèse plus évoluée est la suivante : lumière Voir la circulation des électrons via le schéma en Z chez les plantes actuelles. |
|
5. Chimiosynthèses évoluées (bactéries chimiotrophes) Après l'enrichissement en oxygène de l'atmosphère, des bactéries ont oxydé des minéraux réduits en utilisant l'oxygène comme accepteur final d'électrons. Les électrons arrachés au substrat ont servi à synthétiser des cofacteurs pyridiniques réduits (NADPH) avec synthèse d'ATP. Ces bactéries chimiosynthétiques possédant les enzymes du cycle de Calvin pouvaient réduire le CO2 de l'air. Ces chimiosynthèses évoluées sont des exemples d'autotrophie parfaite. |
|
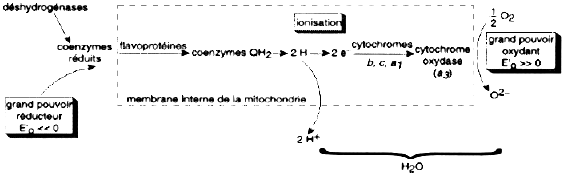
6. Respiration (hétérotrophie évoluée) Finalement, sous une atmosphère riche en oxygène, plusieurs phyla indépendants des bactéries ont utilisé ce gaz comme accepteur final d'électrons lors de l'oxydation composés organiques réduits. Une circulation d'électrons a été établie du substrat respiratoire (acide organique ou cofacteur pyridinique réduit) vers l'oxygène. Ce système permet la récupération de l'énergie libre pour la synthèse d'ATP via la phosphorylation oxydative. Au cours de l'évolution, la chaîne de transporteurs d'électrons utilisée dans les respirations primitives (respiration au nitrate par exemple) se retrouve dans la chaîne respiratoire des bactéries aérobies, l'oxygène se substituant au nitrate comme accepteur final d'électrons.
Source : "Physiologie végétale" (1995) Laval-Martin & Mazliak |
|
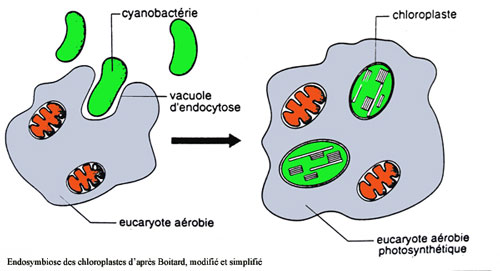
La théorie endosymbiotique de l'origine de la cellule eucaryote postule que la mitochondrie a dérivé d'une bactérie aérobie respirante il y a environ 1,5 milliards d'années. Cette symbiose entre un hôte à métabolisme fermentaire (cytoplasme de la cellule eucaryote) et la mitochondrie (bactérie respirante) illustre le progrès évolutif du passage de l'hétérotrophie primitive anaérobie (fermentation) à l'hétérotrophie aérobie évoluée (respiration). Le bilan énergétique des 2 processus parle de lui-même :
En conclusion, dans une biosphère de plus en plus riche en oxygène, les êtres à métabolisme respiratoire se sont bien mieux développés que les êtres fermentaires.
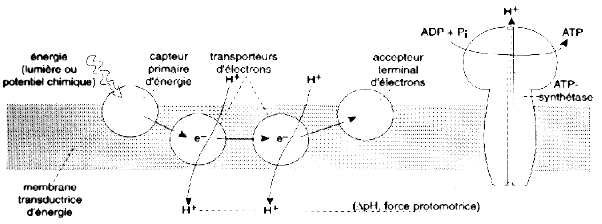
Source : "Physiologie végétale" (1995) Laval-Martin & Mazliak Simultanément à la circulation intramembranaire des électrons s'est mis en place un transfert transmembranaire de protons, créant ainsi une différence de pH de part et d'autre de la membrane. Cela constitue la force "proton motrice" qui est le moteur de l'enzyme qui catalyse la synthèse de l'ATP : l'ATP synthase membranaire. |
![]()