| Promiscuité enzymatique et évolution dirigée des enzymes |
| Tweet |
|
|
1. Introduction 2. Les indices de mesure de la promiscuité enzymatique a. Paramètres cinétiques 3. Facteurs responsables de la promiscuité enzymatique 4. Exemples de promiscuité enzymatique |
a. Divers types de promiscuité 5. La reconstruction (ou résurrection) de protéines ancestrales a. Principe 6. Liens Internet et références bibliographiques |
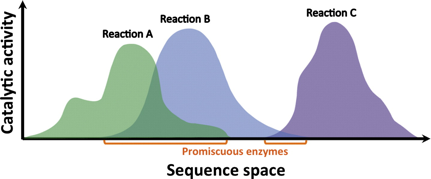
1. Introduction Les enzymes sont des catalyseurs biologiques remarquablement spécifiques. Cependant, de nombreuses enzymes sont capables de catalyser d'autres réactions et/ou de transformer d'autres substrats que celles/ceux pour lesquels elles sont physiologiquement spécialisées : c'est la promiscuité enzymatique ("enzyme promiscuity"). L'un des premiers liens entre promiscuité enzymatique et évolution des protéines a été établi par R.A. Jensen (1976) qui a émis l'hypothèse que les enzymes primordiales (ancestrales), en petit nombre, devaient posséder des spécificités de substrats larges (conférant ainsi de nombreuses capacités métaboliques) contrairement aux enzymes actuelles, pour la plupart hautement spécifiques. Voir un cours sur le monde à ARN. La relation entre l'activité catalytique et l'espace des séquences en acides aminés peut être représentée comme un "paysage catalytique" (pic = activité catalytique élevée et vallée = activité catalytique faible).
Source : Baier & Tokuriki (2014)
On trouve fréquemment le terme anglo-saxon "promiscuous enzyme" que l'on peut traduire par "enzyme promiscueuse".
|
La promiscuité n'est pas réservée aux enzymes : chaque macromolécule dans une cellule peut établir des interactions promiscueuses avec de multiples molécules.
Différence entre promiscuité enzymatique et protéines "moonlighting" La promiscuité enzymatique met en jeu le même site actif pour les différentes activités tandis que les fonctions des protéines "moonlighting" s'appuient sur différents sites de ces protéines. De plus, il est probable que le désordre structural joue un rôle plus important dans la promiscuité enzymatique car elle nécessite qu'un même site de fixation reconnaisse et fixe des substrats très différents. La promiscuité enzymatique est également différente des pseudoenzymes. |
|
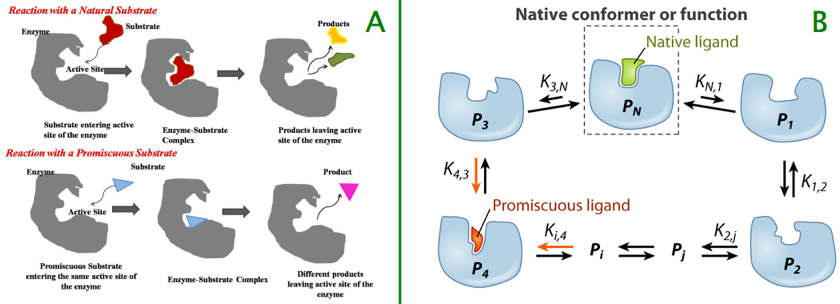
2. Les indices de mesure de la promiscuité enzymatique L'amplitude de la promiscuité enzymatique fait référence aux paramètres cinétiques qui caractérisent l'activité d'une enzyme promiscueuse par rapport à l'activité de l'enzyme native.
Sources : Figure A - Arora et al. (2014); Figure B - Khersonsky & Tawfik (2010) La détoxification des médicaments par certaines enzymes et transporteurs de médicaments illustre bien ces notions. En effet, ces types de protéines doivent reconnaître des substrats d'une grande diversité structurale : les mécanismes de l'évolution ont probablement sélectionné des structures enzymatiques caractérisées par des valeurs de (kcat/KM) en adéquation avec cette large gamme de substrats, plutôt que d'optimiser une valeur de (kcat/KM) pour chaque substrat.
Le paramètre accélération de la vitesse (kcat/knon catalysée) permet d'apprécier l'amplitude des effets de la promiscuité sur les propriétés catalytiques. Le paramètre compétence catalytique [(kcat/KM)/knon catalysée] :
|
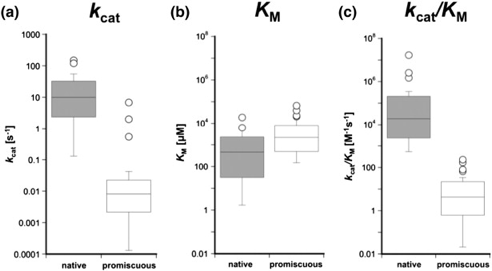
b. Illustration des métallo-β-lactamases Les métallo-β-lactamases (MBL, E.C. 3.5.2.6, hydrolases dépendantes de l'ion mono- ou di-zinc) confèrent une résistance des bactéries à presque tous les antibiotiques β-lactamines. Les membres de la superfamille MBL ont des séquences et des fonctions sensiblement divergentes.
Au moins 24 familles avec des fonctions distinctes ont été identifiées au sein de la superfamille MBL : ces fonctions sont aussi diverses que le traitement de l'ADN, des ARN et des nucléotides, la détection de concentration critique d'autoinducteur dans l'environnement extracellulaire et l'activation de la transcription, la détoxification, la résistance aux antibiotiques, l'hydrolyse des pesticides… Figure ci-dessous : paramètres cinétiques de 20 activités natives et 36 activités promiscueuses de MBL. Les rapports de la médiane des activités natives vs. promiscueuses sont respectivement 1200 pour kcat (10 s-1 vs. 8 10-3 s-1), 5 pour KM (0,46 mM-1 vs. 7,5 mM-1) et 4400 pour le rapport (kcat/KM) (1,9 104 M-1.s-1 vs. 4,3 M-1.s-1).
Source : Baier & Tokuriki (2014) |
3. Facteurs responsables de la promiscuité enzymatique Plusieurs mécanismes moléculaires induisent la promiscuité enzymatique : certains sont intrinsèques au mécanisme de l'enzyme promiscueuse et d'autres sont intrinsèques au partenaire qui interagit avec l'enzyme. L'ajustement induit ("induced fit", IF) et la sélection de conformations ("selected fit" ou "conformational selection", CS ou ) sont 2 modèles qui décrivent la fixation d'un ligand sur une protéine en général (d'un substrat sur une enzyme en particulier). Les enzymes de détoxication promiscueuses :
a. Les modifications post-traductionnelles Par exemple, les modifications par les protéines SUMO. b. Les domaines structuraux multiples Les domaines structuraux permettent aux protéines d'avoir plusieurs fonctions. Ils enrichissent le répertoire fonctionnel d'une superfamille protéique et génèrent des chaînes polypeptidiques qui peuvent accomoder une large gamme de partenaires moléculaires. En raison de leur modularité dans la structure des protéines (donc des enzymes), les domaines peuvent être considérés comme les unités par lesquelles les protéines évoluent. Certains domaines ne sont présents qu'en combinaisons spécifiques. D'autres domaines sont présents dans diverses architectures protéiques et contribuent à leur diversité : ce sont des domaines que l'on peut qualifier de promiscueux. c. L'état d'oligomérisation Il détermine la fonction des protéines. Par exemple, la pyruvate kinase sous forme de tétramère est une enzyme de la glycolyse et sous forme de monomère c'est une protéine de fixation d'hormone thyroïdienne. d. La reconnaissance partielle Des modifications conformationnelles du site de fixation ne sont pas toujours nécessaires pour la promiscuité enzymatique. La reconnaissance moléculaire peut être partielle et résulter d'une complémentarité imparfaite entre l'enzyme et son ligand. La reconnaissance partielle est un mécanisme d'autant plus probable que de nombreuses enzymes catalysent des réactions impliquant une famille de ligands avec des efficacités catalytiques différentes. e. Les sites d'interactions multiples L'existence de plusieurs sites de fixation ou la possibilité de fixer plusieurs ligands sur un seul site sont les principaux facteurs de la promiscuité enzymatique.
f. La spécificité de cofacteurs large
g. La flexibilité structurale et la plasticité du site actif des enzymes C'est le mécanisme le plus important pour rendre compte de la promiscuité enzymatique.
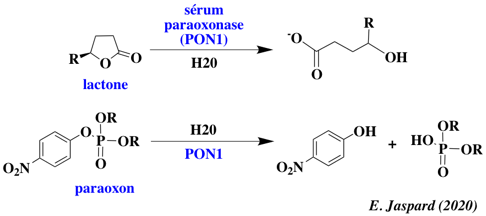
Illustration Il existe plusieurs sous-sites au sein du site actif de certaines enzymes promiscueuses. Par exemple, l'enzyme PON1 ("serum paraoxonase/arylesterase 1") du sérum possède plusieurs activités catalytique :
L'hydrolyse de la lactone (activité native lactonase) est médiée par un mécanisme impliquant une histidine. L'activité promiscueuse phospho-triestérase médiée par d'autres résidus du site actif.
|
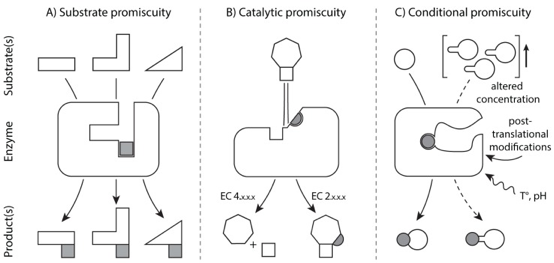
4. Exemples de promiscuité enzymatique a. Divers types de promiscuité Le terme promiscuité a 3 acceptions principales dans la littérature :
Source : Piedrafita et al. (2015) Différentes définitions / classifications de la promiscuité enzymatique : Copley S.D. (2003), Bornscheuer & Kazlauskas (2004), Mariuzza R.A. (2006), Hult & Berglund (2007), Nobeli et al. (2009). |
| Exemple d'enzymes promiscueuses et de réactions | promiscuité de conditions | promiscuité de substrat | promiscuité catalytique |
| réversion de la réaction catalysée par la maltase | oui | - | - |
| réversion de la réaction catalysée par la lipase pancréatique | oui | - | - |
| lipase de foie de porc : réactions dans différents solvants | oui | - | - |
| pyruvate décarboxylase : synthèse de la liaison C-C | - | oui | oui |
| lipase : transacylation en phase gazeuse | oui | - | - |
| subtilisine E : réactions dans différents solvants - mutants | oui | - | - |
| alanine déshydrogénase : utilisation du co-facteur | - | oui | - |
| phosphite déshydrogénase : utilisation du co-facteur | - | oui | - |
| lipase : résolution des acides chiraux | - | oui | - |
| super famille des énolases : activités promiscueuses liées à l'évolution | - | - | oui |
| addition de Markovnicov catalysée par les acylases : formation de la liaison C-N | - | - | oui |
| addition d'aldol catalysée par la lipase : formation de la liaison C-C | - | - | oui |
| formation des liaisons C-O, C-S et C-N catalysée par les sérine hydrolases | - | - | oui |
| addition de conjugués catalysée par les protéases : formation de la liaison C-N | - | - | oui |
| addition de conjugués catalysée par les lipases : formation de la liaison C-N | - | - | oui |
| formation des liaisons C-C, C-S et C-N catalysée par les lipases | - | - | oui |
| anhydrase carbonique en tant que peroxydase | - | - | oui |
| aminopeptidase en tant qu'oxydase | - | - | oui |
| hydrolyse de phosphodiesters par une aminopeptidase | - | - | oui |
| activité aminotransférase catalysée par la racémase dépendante du pyridoxal-phosphate | - | - | oui |
| activité rétro-aldol catalysée par la racémase dépendante du pyridoxal-phosphate | - | - | oui |
| réactions de synthase catalysées par la glycosidase | - | - | oui |
| Source : Hult & Berglund (2007) | |||
b. Autres exemples d'enzymes qui catalysent 2 réactions Enzyme rhamnulose-1-phosphate aldolase (E. coli b3902, rhaD, E.C. 4.1.2.19) :
Enzyme beta-cétoacyl-acyl-carrier-protéine [acp] synthase II (E.C. 2.3.1.179) :
Enzyme bifonctionnelle E.C. 1.5.1.5 (5,10-methylene tétrahydrofolate:NADP+ oxidoreductase) et E.C. 3.5.4.9 (5,10-methenyltetrahydrofolate 5-hydrolase, ring-opening) : |
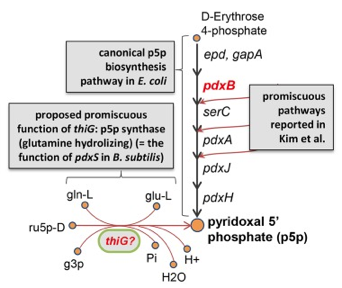
c. Prédiction d'enzymes promiscueuses à partir de modèles métaboliques La méthode GEM-PROPER ("GEnome-scale metabolic Modeling PROmiscuity PrEdictoR") a permis d'identifier les enzymes promiscueuses à partir d'un modèle de reconstruction métabolique à l'échelle du génome de Escherichia Coli.
La paire de gènes [cible-remplaçant] prédits [ΔpdxB/thiG] a été validée expérimentalement chez Escherichia Coli :
Source : Oberhardt et al. (2016) Abréviations : ru5p-D = D-ribulose 5-phosphate; gln-L = L-glutamine; g3p = glycéraldéhyde 3-phosphate; glu-L = L-glutamate; Pi = phosphate. Voir un cours sur les modèles de reconstruction métabolique à l'échelle d'un génome. |
5. La reconstruction (ou résurrection) de protéines ancestrales Il existe deux moyens pour "ressusciter" des molécules biologiques disparues :
La reconstruction de séquences ancestrales ("Ancestral Sequence Reconstruction", ASR) permet d'étudier les séquences et les propriétés des protéines ancestrales.
La résurrection de protéines ancestrales présente un fort potentiel biotechnologique. Voir le projet "Ancestral Sequence Reconstruction Engineering of Coagulation Factors". |
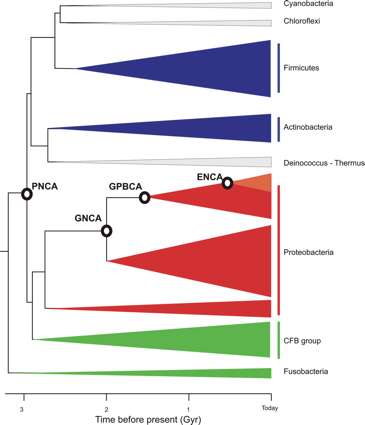
b. Illustration : reconstruction de β-lactamases ancestrales Analyse phylogénique des β-lactamases de classe A Il existe 4 classes (A, B, C et D) de β-lactamases : la classe A (hydrolase à sérine de type pénicillinase) est la plus courante. La phylogénie des séquences de ces β-lactamases a été construite avec une méthode bayésienne :
La phylogénie des β-lactamases de classe A a permis de cibler des noeuds remontant à l'époque du Précambrien (arbre des eubactéries) : dernier ancêtre commun des entérobactéries (ENCA), dernier ancêtre commun des gamma-protéobactéries (GPBCA), dernier ancêtre commun de diverses bactéries Gram- (GNCA) et dernier ancêtre commun de diverses bactéries Gram- et Gram+ (PNCA).
Source : Risso et al. (2013) Synthèse et propriétés structurales et fonctionnelles des β-lactamases ancestrales Ces β-lactamases ancestrales (âge estimé de 2 à 3 milliards d'années) ont été synthétisées puis caractérisées expérimentalement :
Dans le cas des β-lactamases ancestrales, une stabilité élevée et une promiscuité accrue (susceptibles de contribuer à une évolutivité élevée) sont des atouts pour l'étude de l'évolution des protéines et pour élaborer des échafaudages protéiques afin de concevoir de nouvelles molécules. |
| 6. Liens Internet et références bibliographiques |
Structure-Function Linkage Database Binding Mother of All Databases ProtASR : an evolutionary framework to reconstruct ancestral protein sequences accounting for structural constraints PhyloBot: a web portal for automated phylogenetics, ancestral sequence reconstruction, and exploration of mutational trajectories |
|
Pocker & Stone (1965) "The catalytic versatility of erythrocyte carbonic anhydrase. The enzyme - catalyzed hydrolysis of rhonitrophenyl acetate" J. Am. Chem. Soc. 87, 5497 - 5498 Jensen R.A. (1976) "Enzyme recruitment in evolution of new function" Annu. Rev. Microbiol. 30, 409 - 425 O'Brien & Herschlag (1999) "Catalytic promiscuity and the evolution of new enzymatic activities" Chem. Biol. 6, R91 - R105 |
|
Hult & Berglund (2007) "Enzyme promiscuity: mechanism and applications" Trends Biotechnol. 25, 231 - 238 Johnson K.A. (2008) "Role of induced fit in enzyme specificity : a molecular forward/reverse switch" J Biol Chem. 283, 26297 - 26301 Nobeli et al. (2009) "Protein promiscuity and its implications for biotechnology" Nat. Biotechnol. 27, 157 - 167 |
|
Khersonsky & Tawfik (2010) "Enzyme promiscuity: a mechanistic and evolutionary perspective" Annu. Rev. Biochem. 79, 471 - 505 Carbonell & Faulon (2010) "Molecular signatures-based prediction of enzyme promiscuity" Bioinformatics 26, 2012 - 2019 Khersonsky & Tawfik (2010) "Enzyme promiscuity : a mechanistic and evolutionary perspective" Annu. Rev. Biochem. 79, 471 - 505 |
|
Kim et al. (2013) "NDM-1, the ultimate promiscuous enzyme: substrate recognition and catalytic mechanism" FASEB J. 27, 1917 - 1927 Jia et al. (2013) "Multifunctional enzymes in archaea: promiscuity and moonlight" Extremophiles 17, 193 - 203 Gao & Skolnick (2013) "A comprehensive survey of small-molecule binding pockets in proteins" PLoS Comput. Biol. 9, e1003302 Risso et al. (2013) "Hyperstability and substrate promiscuity in laboratory resurrections of Precambrian β-lactamases" J. Am. Chem. Soc. 135, 2899 - 2902 |
|
Arora et al. (2014) "Enzyme promiscuity: using the dark side of enzyme specificity in white biotechnology" Sustain. Chem. Proc. 2, 25 Baier & Tokuriki (2014) "Connectivity between catalytic landscapes of the metallo-β-lactamase superfamily" J. Mol. Biol. 426, 2442 - 2456 Kaltenbach & Tokuriki (2014) "Dynamics and constraints of enzyme evolution" J. Exp. Zool. B Mol. Dev. Evol. 322, 468 - 487 |
|
Copley S.D. (2015) "An evolutionary biochemist's perspective on promiscuity" Trends Biochem. Sci. 40, 72 - 78 Piedrafita et al. (2015) "The Impact of Non-Enzymatic Reactions and Enzyme Promiscuity on Cellular Metabolism during (Oxidative) Stress Conditions" Biomolecules 5, 2101 - 2122 |
|
Clifton & Jackson (2016) "Ancestral protein reconstruction yields insights into adaptive evolution of binding specificity in solute-binding proteins" Cell. Chem. Biol. 23, 236 - 245 Merkl & Sterner (2016) "Ancestral protein reconstruction: techniques and applications" Biol. Chem. 397, 1 - 21 Patrick et al. (2016) "Biochemical characterization of predicted Precambrian RuBisCO" Nat. Commun. 7, 10382 Oberhardt et al. (2016) "Systems-Wide Prediction of Enzyme Promiscuity Reveals a New Underground Alternative Route for Pyridoxal 5'-Phosphate Production in E. coli" PLoS Comput. Biol. 12, e1004705 |
|
Chen et al. (2019) "Promiscuous enzymatic activity-aided multiple-pathway network design for metabolic flux rearrangement in hydroxytyrosol biosynthesis" Nat. Commun. 10, 960 Atkins W.M. (2019) "Mechanisms of promiscuity among drug metabolizing enzymes and drug transporters" FEBS J. doi: 10.1111/febs.15116 Cerisier et al. (2019) "High impact : the role of promiscuous binding sites in polypharmacology" Molecules 24, 2529 van Loo et al. (2019) "Transition-state interactions in a promiscuous enzyme: sulfate and phosphate monoester hydrolysis by Pseudomonas aeruginosa arylsulfatase" Biochemistry 58, 1363 - 1378 |
![]()