Métabolisme mitochondrial lié à la chaîne respiratoire |
|
1. La mitochondrie et la chaîne respiratoire 2. Caractéristiques générales des complexes protéiques de la chaîne respiratoire 3. Rôles biologiques des formes réduites NADH et NADPH 4. Les systèmes de navette pour transporter le pouvoir réducteur a. La navette glycérol phosphate 5. La force proton-motrice et la synthèse d'ATP par l'ATP synthase 6. Transport du Pi, de l'ADP et de l'ATP a. Transport du Pi au travers de la membrane interne |
d. Transport de l'ATP au travers de la membrane externe 7. Découplage [dissipation du gradient de protons / synthèse d'ATP] a. Les protéines de découplage 8. Les espèces réactives de l'oxygène de la mitochondrie a. Généralités 9. Quelques inhibiteurs et agents découplants 10. Liens Internet et références bibliographiques |
|
|
| 1. La mitochondrie et la chaîne
respiratoire
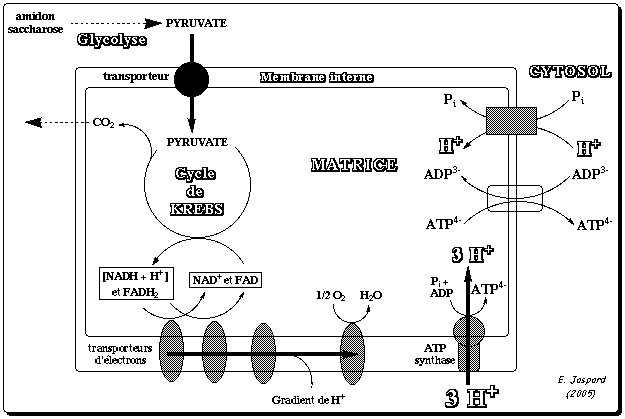
La mitochondrie est limitée par deux membranes caractérisées par des propriétés très différentes : La membrane externe est pauvre en protéines. Elle contient, en particulier, des protéines transmembranaires, les porines, qui permettent le passage des ions et des métabolites hydrosolubles de masse molaire < 7000 Daltons. A l'inverse, la membrane interne est très riche en protéines mais elle est quasiment imperméable aux ions et aux métabolites hydrosolubles. Ces substances ne peuvent traverser la membrane qu'à l'aide de protéines membranaires de transport (l'ATP, l'ADP et le Pi sont transportés par ce type de protéines), ou de systèmes plus complexes qu'on appelle navette. L'espace entre ces deux membranes s'appelle l'espace intermembranaire. La zone interne de la mitochondrie (bordée par la membrane interne) s'appelle la matrice. Elle contient les enzymes du cycle de Krebs et la plupart de celles qui catalysent l'oxydation des acides gras. La chaîne respiratoire est localisée dans la membrane interne des mitochondries. Le nombre des crêtes accroit la surface de cette membrane et ainsi chaque mitochondrie contient des milliers d'exemplaires de la chaîne de transport d'électrons. Les crêtes pénètrent dans la matrice. Certaines protéines mitochondriales sont synthétisées par la mitochondrie (génome mitochondrial), mais la plupart d'entre elles sont codées par le génome nucléaire puis importées dans la mitochondrie. Voir un cours sur la biogénèse (synthèse, adressage et assemblage) des protéines membranaires intégrales. |
| 2.
Caractéristiques générales des complexes protéiques de la chaîne respiratoire
La chaîne respiratoire est un ensemble de complexes protéiques qui assurent un transfert de protons et/ou d'électrons. Les caractéristiques des complexes protéiques sont les suivants : |
| Règne | Complexe ou protéine | Sous-unités | Masse molaire | Composants | Transfert | Inhibiteurs |
| Animaux et végétaux | NADH - coenzyme Q |
25 | 800 kDa | 1 FMN 22 à 24 atomes [Fe - S] dans 5 à 8 centres | électrons protons |
roténone amytal |
| succinate
- coenzyme Q oxydoréductase (complexe II) |
4 | 125 kDa | 1 FAD 7 à 8 atomes Fe - S dans 3 centres cytochrome b560 | électrons | malonate | |
| coenzyme Q | 1 | 0,86 | lipide isoprénoide | électrons protons |
----- | |
| Spécifiques des végétaux | NAD(P)H déshydrogénases | 4 protéines : 2 NDint et 2 NDext cytochrome b560 | électrons | diphénylèneiodonium (DPI) insensibles à la roténone | ||
| oxydase alternative (AOX) | 2 | 64 kDa | électrons | SHAM propylgallate insensible au KCN | ||
| Animaux et végétaux | coenzyme Q
- cytochrome c oxydoréductase (complexe III) |
8 | 220 kDa | 2 centres [Fe - S] cytochrome b560 cytochrome b566 cytochrome c1 | électrons protons |
antimycine |
| cytochrome C | --- | 12 kDa | hème (Fe) | électron | ----- | |
| cytochrome c oxydase (complexe IV) |
12 | 200 kDa | cytochrome a cytochrome a3 2 ions cuivre |
électrons protons |
KCN / CO / NaN3 |
3. Rôles biologiques des formes réduites NADH et NADPH Malgré des structures très semblables, les formes réduites de ces deux coenzymes sont employées dans la cellule de manière très différente. Le NADH est produit par des réactions de voies du catabolisme (dégradation) :
Le NADPH est produit par des réactions de voies de l'anabolisme (biosynthèse) telles que la voie des pentoses phosphate.
|
|
4. Les systèmes de navette ("shuttle") pour transporter le pouvoir réducteur Au cours de la glycolyse, 2 molécules de NAD réduit (NADH) sont formées par molécule de glucose dégradé.
|
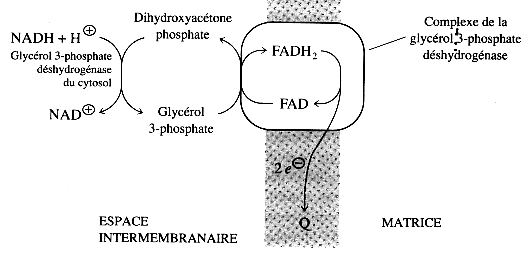
a. La navette glycérol phosphate Elle met en jeu 2 enzymes :
Le pouvoir réducteur entre dans la mitochondrie de la manière suivante :
|
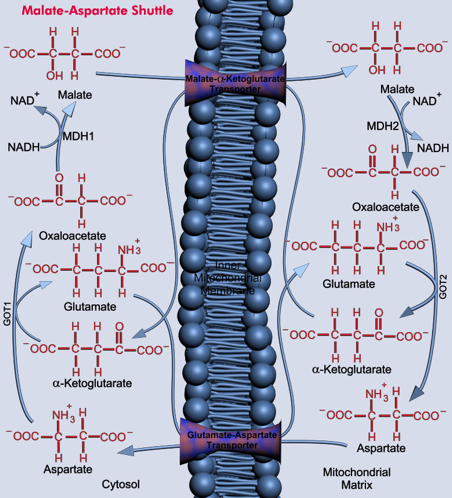
b. La navette malate - aspartate Elle met en jeu de nombreuses enzymes et transporteurs membranaires : en particulier, la malate déshydrogénase (MDH1 et MDH2 - E.C. 1.1.1.37) et l'aspartate transaminase (GOT1 et GOT2, E.C. 2.6.1.1), présentes à la fois dans le cytosol et dans la mitochondrie. Les principales étapes du mécanisme sophistiqué par lequel le pouvoir réducteur du NADH entre dans la mitochondrie sont les suivantes :
En conséquence, 3 molécules d'ATP sont synthétisées lors de la réoxydation du NADH formé au cours de la glycolyse quand le pouvoir réducteur entre dans la mitochondrie via la navette malate - aspartate. |
5. La force proton-motrice et la synthèse d'ATP par l'ATP synthase La théorie chimio-osmotique formulée par Peter Mitchell en 1961 (Prix Nobel en 1978) postule que :
|
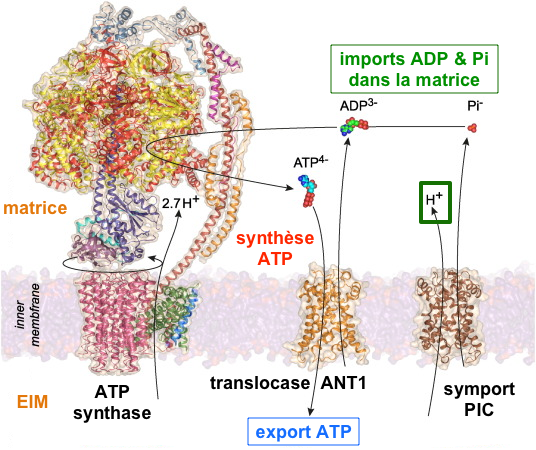
6. Transport du Pi, de l'ADP et de l'ATP a. Transport du Pi au travers de la membrane interne Le phosphate inorganique Pi est importé de l'espace intermembranaire vers la matrice par le transporteur du Pi ("Phosphate Carrier" - PiC) ou SLC25A3 ("Solute Carrier family 25 member 3").
Figure adaptée de : Kunji et al. (2016) PiC possède 6 domaines transmembranaires et ses extrémités N- et C-terminales se trouvent dans la matrice. Sa structure est similaire à ANT1 et UCP1. Pi est utilisé pour :
|
b. Transport de l'ADP et de l'ATP au travers de la membrane interne La quantité d'ATP nécessaire aux besoins de la cellule hors mitochondrie est exportée de la matrice vers l'espace intermembranaire par un antiport qui transporte l'ADP en sens inverse : la translocase [ADP/ATP] 1 (SLC25A4 - "Solute Carrier family 25 member 4") ou ANT1. C'est l'une des protéines les plus abondantes de la membrane interne des mitochondries qui assure le premier transport et le dernier transport (import d'ADP / export d'ATP) de la phosphorylation oxydative des eucaryotes. On estime que chaque jour, l'équivalent du poids du corps humain en ADP et en ATP est transporté par l'ensemble des translocases des mitochondries de toutes les cellules pour assurer les processus cellulaires nécessitant de l'énergie. La chaîne polypeptidique de la translocase ANT1 :
Fonctionnement de la translocase ANT1 C'est un antiport : ADP3-espace intermembranaire + ATP4-matrice <=> ADP3-matrice + ATP4-espace intermembranaire
ANT1 change de conformations entre un état ouvert dans le cytoplasme (état c) et un état ouvert dans la matrice (état m) avec une fixation alternée du ligand sur un seul site orienté alternativement vers le cytosol et vers la matrice. |
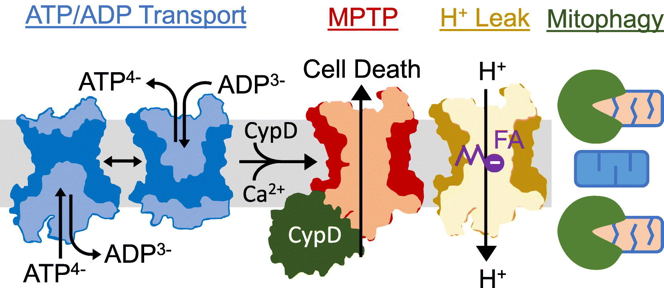
c. Autres fonctions de la translocase ANT1 ANT1 assure également le transport de protons : cette fonction contribue au découplage [chaîne de transport d'électrons / ATP synthase] qui provoque la thermogenèse mitochondriale. H+espace intermembranaire <=>H+matrice L'activité transport de protons (thermogénèse) de ANT1 est inhibée par son activité antiport [ADP:ATP] (synthèse d'ATP) : ANT1 (SLC25A4) participe donc à la régulation de production d'énergie dans la mitochondrie en maintenant un équilibre entre ces 2 activités opposées.
Source : Bround et al. (2020) ANT1 a également un rôle important dans l'ouverture des pores de transition de perméabilité mitochondriale ("Mitochondrial Permeability Transition Pore" - MPTP) de la membrane interne qui s'ouvrent en réponse à une forte élévation de la concentration du Ca2+ dans la matrice : l'organite gonfle puis rompt, provoquant la mort cellulaire. Enfin, ANT1 favorise la mitophagie via son interaction avec TIM44 : la translocase de préséquence TIMM23 est inhibée, ce qui contribue à stabiliser PINK1. |
d. Transport de l'ATP au travers de la membrane externe Le canal sélectif d'anions dépendant du potentiel de membrane ("Voltage-Dependent Anion-selective Channel" - VDAC) est une protéine membranaire en tonneau β ("β-barrel") de la membrane externe ("outer membrane") de la mitochondrie qui régule l'entrée / sortie de métabolites et d'ions :
VDAC adopte une conformation :
Voir un cours sur les transports. Autres rôles de VDAC Des métabolites tels que NADH et ATP établissent un lien entre la glycolyse (cytosol) et la phosphorylation oxydative (membrane interne de la mitochondrie). En conséquence, le rôle essentiel de VDAC dans le métabolisme énergétique et dans l'homéostasie du calcium en fait une cible thérapeutique importante. Par ailleurs, VDAC participe à la réponse au stress de la mitochondrie en régulant la formation des pores MPTP (voir ci-dessus) formés dans la membrane interne lors d'un fort stress oxydatif ou d'un taux de calcium élevé dans le cytosol. |
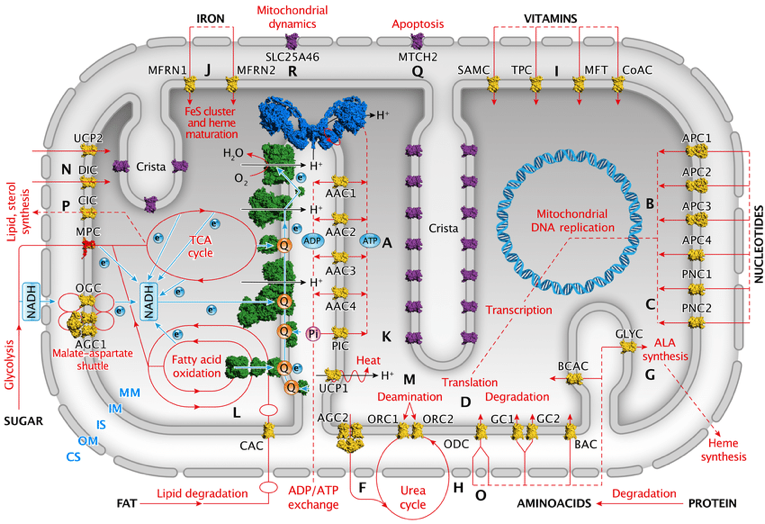
e. Vision globale des transporteurs membranaires de la mitochondrie Les complexes de la chaîne de transport d'électrons sont représentés en vert et l'ATP synthase est en bleu (au centre de la figure).
Source : Kunji et al. (2020) Q : ubiquinone ; Pi : phosphate; ALA : acide aminolévulinique. Les protéines de transport identifiées sont en jaune et celles qui ne sont pas identifiées sont en violet.
|
7. Découplage [dissipation du gradient de protons / synthèse d'ATP] a. Les protéines de découplage Cinq isoformes de protéine de découplage ("UnCoupling Protein" - UCP) sont identifiées chez les mammifères (UCP1 à UCP5).
|
|
La protéine de découplage 1 ("Mitochondrial brown fat uncoupling protein 1" - UCP1) se trouve principalement dans les mitochondries du tissu adipeux brun des mammifères. UCP1 est un symport ("Solute Carrier family 25 member 7" - SLC25A7) qui transporte un acide gras à longue chaîne (AGLC) et un proton. UCP1 est responsable de la perméabilité aux protons de la membrane interne des mitochondries de ce tissu.
Les adipocytes bruns sont donc impliqués dans la production de chaleur lors de la thermogenèse dite "sans frisson" (adaptée aux variations de température et d'alimentation). |
|
L'activité d'UCP1 :
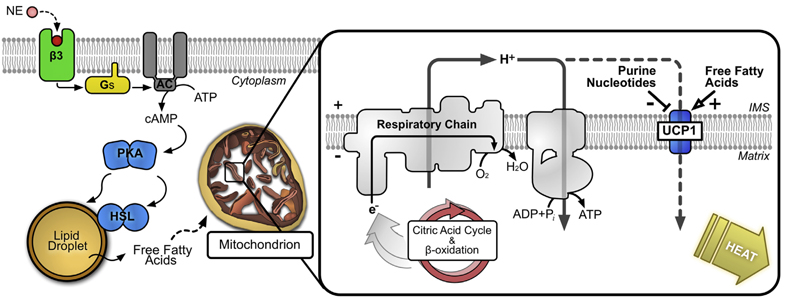
Ces derniers sont produits dans les adipocytes bruns par la lipolyse de gouttelettes lipidiques issues du cytoplasme lors d'une stimulation adrénergique.
Source : Crichton et al. (2017) Partie gauche de la figure : la noradrénaline (NE) libérée en réponse à un stimulus physiologique active une voie dépendante de l'AMP cyclique (AMPc) qui libère des acides gras à partir des réserves lipidiques cellulaires. Partie droite de la figure :
Il existe au moins 3 protéines de découplage mitochondriales qui participent à des fonctions spécifiques : la thermogenèse "sans frisson" (UCP1), la signalisation de la satiété (UCP2) et le métabolisme musculaire (UCP3). |
|
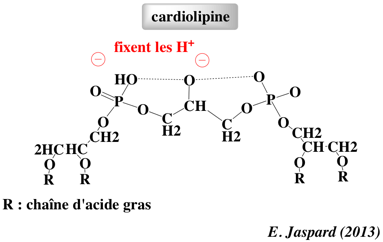
UCP1 fixe 3 molécules de cardiolipine qui stabilisent sa structure. La cardiolipine (diphosphatidylglycérol) est un glycérophospholipide qui représente 18 à 20% des phospholipides de la membrane interne des mitochondries.
La cardiolipine a de nombreux rôles :
|
8. Les espèces réactives de l'oxygène de la mitochondrie Les espèces réactives de l'oxygène ("Reactive Oxygen Species" - ROS) sont des dérivés de l'oxygène générés par le métabolisme cellulaire de toutes cellules qui utilise l'oxygène.
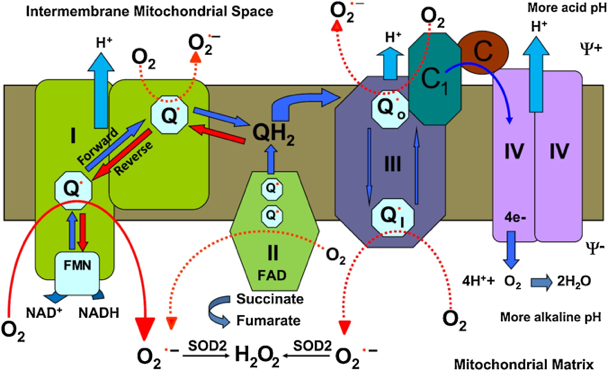
Les ROS mitochondriales ("mitochondrial ROS" - mtROS) sont générées par les mitochondries.
Source : Koju et al. (2019) Le superoxyde est transformé en H2O2 par des superoxyde dismutases (oxydoréductases) : 2 H+ + 2 O2°- <=> H2O2 + O2 La superoxyde dismutase 1 (SOD1 - E.C. 1.15.1.1) agit dans l'espace intermembranaire et la superoxyde dismutase 2 (SOD2) agit dans la matrice. |
|
Avantages Les mtROS ont un rôle important de molécules de signalisation. Exemples : la prolifération cellulaire, l'adaptation à l'hypoxie, la détermination du devenir cellulaire, la signalisation antivirale mitochondriale et certaines réponses antivirales, ... Les ROS sont impliquées dans différentes cascades de signalisation des protéines kinases. Exemples : les voies de la protéine kinase B, de la protéine kinase activée par l'AMP (AMPK) et de la PK3 activée par le mitogène qui agit sur le devenir de la cellule (autophagie vs. apoptose). Les mtROS sont des composants antimicrobiens impliqués dans la défense innée contre les infections bactériennes. Inconvénients Un excès de ROS peut provoquer des dommages cellulaires irréversibles, voire la mort cellulaire. La survenue de nombreuses maladies et l'hypoxie sont étroitement liées à l'augmentation de la production de ROS. |
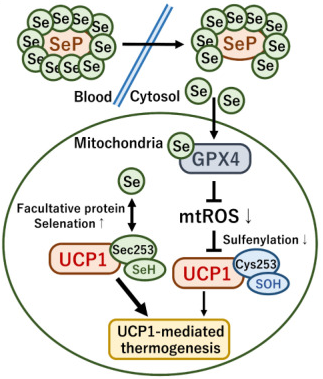
c. Sélénium : UCP1, séléno-cystéine et ROS Le sélénium (34Se) est impliqué dans le métabolisme énergétique des mitochondries et dans la capacité thermogénique du tissu adipeux.
Figure ci-dessous, à gauche : la sélénocystéine (Sec) peut être incorporée de manière facultative dans UCP1 (Jedrychowski et al., 2020).
La supplémentation alimentaire en sélénium :
Source : Takeda et al. (2023) Figure ci-dessus à droite : le sélénium délivré par les SeP dans les mitochondries active la glutathion peroxydase 4 (GPX4), enzyme anti-oxydante dépendante du sélénium. En conséquence :
|
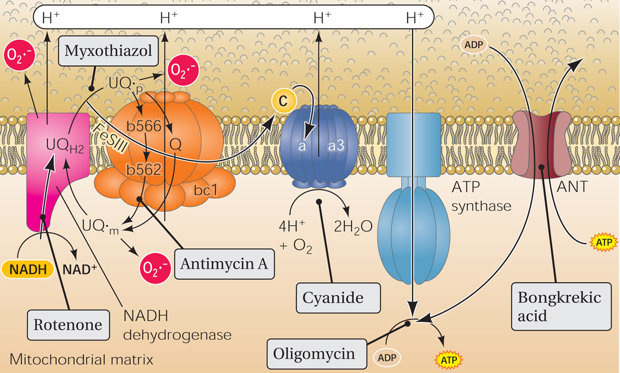
9. Quelques inhibiteurs et agents découplants des processus de la respiration
Source : O'Rourke et al. (2005) |
| Processus ou complexe inhibés | Inhibiteur |
| Inhibiteurs de la phosphorylation qui empêchent la consommation d'oxygène après l'addition d'ADP mais n'ont pas d'effet sur la stimulation de la respiration par les agents découplants. | oligomycine |
Agents découplants
|
2,4 dinitrophénol carbonyl cyanide m-chloro-phenyl hydrazone (CCCP) p-trifluoromethoxy-carbonyl-cyanide-phenyl hydrazone (FCCP) |
| Inhibition de l'antiport DIC ("DiCarboxylate Carrier" - SLC25A10) qui échange des acides dicarboxyliques (par exemple, le malate ou le succinate) contre le Pi. | butylmalonate |
| Inhibition du transport de l'ADP ou de l'ATP (translocase ANT1) au travers de la membrane interne (empêche l'exportation de l'ATP ou l'importation de l'ADP). La fixation de l'atractyloside et du BKA ont des effets opposés sur l'efflux des nucléotides par la translocase. |
atractyloside (glycoside diterpénoïde) N-éthyl maléimide acide bongkrekique (BKA) carboxy-atractylate |
| Flux de protons médié par UCP1. | inhibiteurs : ATP, ADP, GTP et GDP activateurs : ROS et acides gras |
| Ionophores qui rendent la membrane interne perméable à des composés qui normalement ne peuvent pas la traverser. | valinomycine, nigericine |
| NADH - coenzyme Q oxydoréductase (complexe I) | roténone |
| succinate - coenzyme Q oxydoréductase (complexe II) | acide malonique |
| coenzyme Q - cytochrome c oxydoréductase (complexe III) | antimycine A |
| cytochrome c oxydase (complexe IV) | KCN (cyanure de potassium) / CO / NaN3 (azoture de sodium) |
| Pour plus de détails, aller au site "Uncouplers and Inhibitors". | |
| 10. Liens Internet et références bibliographiques |
| "Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X | |
Nicholls & Locke (1984) « Thermogenic mechanisms in brown fat » Physiol. Rev. 64, 1 - 64 O'Rourke et al. (2005) "Mitochondrial Ion Channels: Gatekeepers of Life and Death" Physiology (Bethesda) 20, 303 - 315 |
|
|
Kunji et al. (2016) "The transport mechanism of the mitochondrial ADP/ATP carrier" Biochim. Biophys. Acta 1863, 2379 - 2393 Crichton et al. (2017) "The molecular features of uncoupling protein 1 support a conventional mitochondrial carrier-like mechanism" Biochimie 134, 35 - 50 Xiao et al. (2018) "NAD(H) and NADP(H) Redox Couples and Cellular Energy Metabolism" Antioxid. Redox Signal. 28, 251 - 272 |
|
Koju et al. (2019) "Pharmacological strategies to lower crosstalk between nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and mitochondria" Biomed. Pharmacother. 111, 1478 - 1498 Zhao et al. (2019) "Mitochondrial electron transport chain, ROS generation and uncoupling" Int. J. Mol. Med. 44, 3 - 15 |
|
Kunji et al. (2020) "The SLC25 Carrier Family: Important Transport Proteins in Mitochondrial Physiology and Pathology" Physiology (Bethesda) 35, 302 - 327 Bround et al. (2020) "A 20/20 view of ANT function in mitochondrial biology and necrotic cell death" J. Mol. Cell. Cardiol. 144, A3 - A13 Jedrychowski et al. (2020) "Facultative protein selenation regulates redox sensitivity, adipose tissue thermogenesis, and obesity" Proc. Natl. Acad. Sci. 117, 10789 - 10796 |
|
| Takeda et al. (2023) "Mitochondrial Energy Metabolism in the Regulation of Thermogenic Brown Fats and Human Metabolic Diseases" Int. J. Mol. Sci. 24, 1352 | |