Optimisation des conditions d'expression et purification partielle de la forme mature de la protéine de choc thermique HSP22 recombinante de pois (Pisum sativum) Matériels et méthodes |
Voir un cours sur les protéines de stress LEA et HSP. |
1. Caractéristiques des souches d'Escherichia coli 2. La construction dans le vecteur d'expression pET-3a 3. Culture des souches et induction de l'expression 4. Purification partielle de l'HSP22 recombinante |
1. Caractéristiques des souches d'Escherichia coli a. Les deux souches utilisées, BL21(DE3) et B834(DE3)pLysS, sont des dérivés lysogènes de la souche classique E. coli B : elles contiennent le prophage λ DE3 dont l'un des gènes code l'ARN polymérase du bactériophage T7, sous le contrôle du promoteur lacUV5 inductible par l'isopropyl β-D-1-thiogalactopyranoside (IPTG). Ainsi, quand ces souches sont transformées par un vecteur d'expression (comme ceux de la famille pET) qui contient un promoteur fort (celui du bactériophage T7), la synthèse d'ARNm est accrue et la synthèse de la protéine d'intérêt peut atteindre une proportion de 40% à 50% des protéines totales (Baneyx, 1999). Cependant, dans le cas de promoteurs forts, il y a une expression de base en absence d'inducteur. Cette " fuite " peut avoir 2 conséquences :
L'une des stratégies adoptées pour palier ces problèmes est la co-sur-expression du lysozyme du bactériophage T7, dont le gène est porté par un plasmide additionnel, pLysS. En effet, le lysozyme est un inhibiteur de l'ARN polymérase du bactériophage T7, ce qui diminue le niveau de base de la transcription. De plus, l'attaque du peptidoglycane de la membrane externe de E. coli par le lysozyme est très limitée car celui-ci ne peut pas franchir la membrane interne de la bactérie. b. Une autre caractéristique importante de ces deux souches (et de toute souche dérivée de BL21) est qu'elles ne possèdent pas les gènes ompT et lon qui codent pour des protéases qui dégraderaient la protéine sur-exprimée. c. La souche B834 est une souche parentale de BL21, auxotrophe pour la méthionine. |
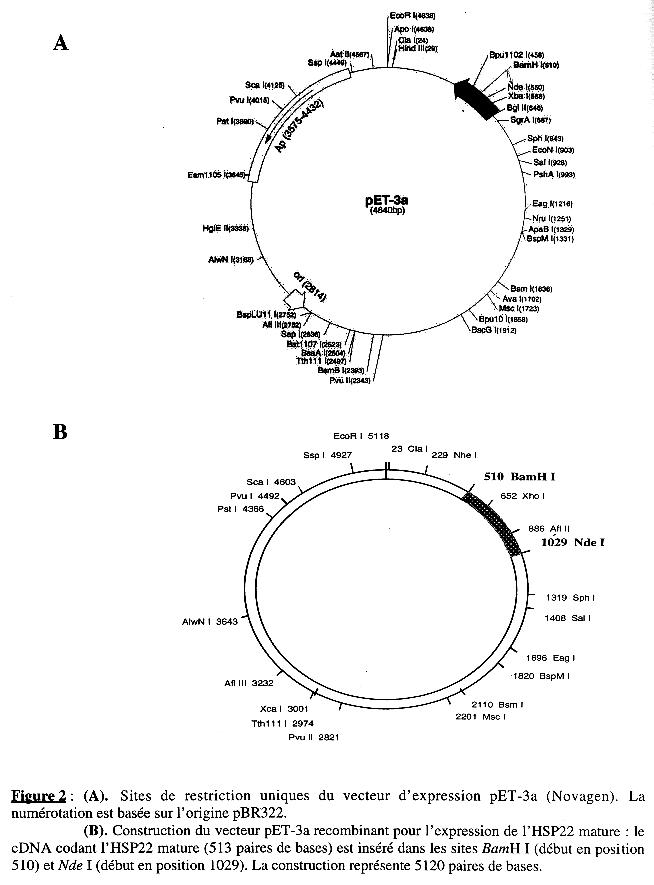
2. La construction dans le vecteur d'expression pET-3a a. La séquence nucléotidique d'origine La séquence qui code pour le précurseur de l'HSP22 de mitochondries de feuilles de pois présente les caractéristiques suivantes (Figure 1) :
|
b. La séquence nucléotidique clonée La séquence codant la forme mature de l'HSP22 a été clonée dans le vecteur pET-3a (Figure 2A), entre les sites NdeI et BamHI (Figure 2B) :
La présence de l'insert et sa taille ont été vérifiées par amplification PCR à partir d'une colonie isolée de chaque souche transformée (voir le paragraphe "3. Culture des souches"). Les séquences des amorces utilisées sont les suivantes :
L'amplification a été effectuée dans le mélange suivant (50 µL totaux) :
L'amplification (30 cycles) a été effectuée dans un thermocycleur programmé de la manière suivante : 1 minute à 94°C; 1 minute à 53°C (pour BL21(DE3)) ou à 50°C (pour B834(DE3)pLysS); 1 minute à 72°C. Les produits d'amplification ont été précipités (isopropanol et acétate d'ammonium). Les culots ont été lavés à l'éthanol et repris dans 15 µL de Tris-EDTA (10:1) puis déposés sur un gel d'agarose 1%. Le profil obtenu après coloration du gel au bromure d'éthidium est présenté dans la Figure 3. La taille du produit d'amplification (537 pb) correspond à celle attendue pour la séquence clonée (513 pb pour la séquence codant plus 24 pb pour les deux sites de restriction et les extensions 5' et 3') dans le vecteur pET-3a. Dans le cas de BL21(DE3), aucun produit d'amplification n'est obtenu pour une température d'hybridation de 50°C. C'est la raison pour laquelle la température d'hybridation utilisée est de 53°C. Cependant, une bande d'amplification parasite est observée (Figure 3A) et cette bande persiste pour une température d'hybridation encore plus élevée (55°C, résultat non présenté). |
c. Les séquences polypeptidiques L'HSP22 précurseur est une protéine de 202 acides aminés d'une masse molaire apparente de 22,9 kDa, dont les caractéristiques sont les suivantes (Figure 1) :
La protéine sur-exprimée (la forme mature de l'HSP22) contient 170 acides aminés pour une masse molaire prédite de 19,5 kDa. L'extrémité N-terminale de la protéine purifiée à partir des mitochondries est l'Asn 33 (Lenne & Douce, 1994). Une analyse par dégradation d'Edmann de l'extrémité N-terminale de la protéine recombinante a mis en évidence l'existence d'une méthionine additionnelle avant l'asparagine 33. |
3. Culture des souches et induction de l'expression Les souches de E. coli ont été cultivées sur milieu "Lysogeny Broth" (LB, Luria Bertani), solide ou liquide, en présence d'antibiotique(s) : carbénicilline seule (200 µg/mL, concentration finale) pour BL21(DE3) et carbénicilline plus chloramphénicol (34 µg/mL) pour B834(DE3)pLysS, la résistance au chloramphénicol étant conférée par le plasmide pLysS. La composition du milieu LB est la suivante (pour 1 litre) : bactotryptone 10 g ; extrait de levure 5 g ; NaCl 10 g et agarose 15 g pour les cultures sur boîte de Pétri. Le milieu a été autoclavé (121°C, 20 min) puis les antibiotiques ont été ajoutés après filtration sur membrane 0,2 µm. Pour chaque expérience (hormis la purification de l'HSP22), le protocole de pré-culture, de culture et d'induction de la transcription du gène codant la protéine recombinante a été le suivant: a. pré-culture sur boîte Les souches ont été repiquées sur une boîte contenant du milieu frais (avec les antibiotiques adaptés) et cultivées une nuit à 30°C. A cette température, les bactéries poussent moins rapidement ce qui permet l'obtention de colonies isolées et également ce qui minimise les risques d' "expulsion" du plasmide recombinant. b. pré-culture liquide Le lendemain, une pré-culture a été effectuée à 37°C à partir d'une colonie isolée dans 10 mL de milieu LB avec les antibiotiques jusqu'à une densité optique A600 = 0,6 à 0,8. Puis 1 ml de cette pré-culture a été prélevé et centifugé (15 min, 4000 rpm). Le culot de bactérie a été repris dans 1 mL de milieu LB frais. Quand l'induction de l'expression de la protéine recombinante ne pouvait être faite le jour même, le culot re-suspendu a été laissé une nuit au frigidaire. c. induction de l'expression 10 mL de LB, contenant divers additifs (tableau ci-après), ont été ensemencés avec 200 µL de culot re-suspendu jusqu'à une densité optique A600 = 0,4 à 0,6. L'expression de la protéine recombinante a alors été induite par addition d'IPTG, dans diverses conditions :
Le témoin de chaque expérience a été cultivé à 37°C, mais sans additif et l'expression a été induite par [IPTG] 0,4 mM. d. prélèvement et analyse des échantillons Deux fois 1 ml de chaque culture ont été prélevés immédiatement (témoin 0 heure d'induction) et après des temps variables d'induction et la densité optique à 600 nm a été mesurée. Ces 2 mL ont été centrifugés (15 min, 4000 rpm, centrifugeuse de paillasse) et le culot a été repris dans 100 µL de Tris-HCl 0,1 M, pH 8. Le contenu protéique de ce lysat brut a été analysé par électrophorèse sur gel d'acrylamide (11%) en conditions dénaturantes (Laemmli, 1970). Selon l'expérience, les dépôts étaient de volume identique ou de quantité de protéines identique pour tous les échantillons. La concentration en protéines totales a été déterminée par la méthode de Bradford (1976).
|
a. Culture et lyse des bactéries Les souches ont été repiquées sur une boîte fraîche (avec les antibiotiques adaptés) et cultivées une nuit à 30°C.
L'expression de la protéine recombinante a été induite par l'addition d'isopropyl β-D-1-thiogalactopyranoside (IPTG) 0,4 mM (concentration finale) quand la culture a atteint une densité optique A600 = 0,44 et 0,41 pour BL21(DE3) et B834(DE3)pLysS, respectivement. L'induction de l'expression a duré 20 heures. Les bactéries ont été centrifugées 15 min à 6000g. Le culot bactérien a été repris dans 10 mL de LB frais, et la même centrifugation a été effectuée. Le culot bactérien a été resuspendu dans 7 mL de tampon de lyse constitué de : Tris-HCl 50 mM, pH 7,5 ; EDTA 1 mM ; PMSF 1 mM ; NaCl 100 mM ; sarcosine 0,2% ; lysozyme 0,25 mg/mL ; DNase I 60 unités, et incubé 2 heures à 37°C. Le lysat a été centrifugé 40 min à 15000 g. Les composants du milieu de lyse ont pour but de déstructurer la membrane bactérienne : (i) l'EDTA, chélateur de
cations divalents, inhibe les métalloprotéases et joue aussi
un rôle d'antioxydant |
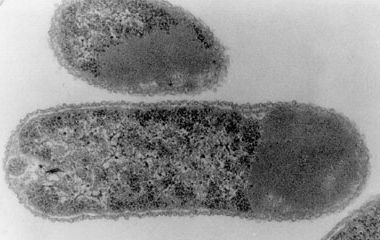
b. Préparation des corps d'inclusion Une conséquence de la transformation de bactéries par un vecteur d'expression qui possède un promoteur fort, mais également du caractère réducteur du cytoplasme (le rapport glutathion forme réduite / forme oxydée est estimé entre 30:1 et 100:1, chez les eucaryotes), est l'accumulation de certaines protéines recombinantes dans des corps d'inclusion (figure ci-dessous) :
Source : Hauke et al. (1998) Curr. Opin. Biotechnol. 9, 497 - 501 Partant du principe que la forte densité de ces particules permet de les isoler par centrifugation, les corps d'inclusion ont été préparés de la manière suivante :
|
c. Solubilisation des agrégats protéiques Les corps d'inclusion sont constitués d'agrégats de la protéine recombinante (elle est donc dans une forme dénaturée) entourés par une membrane : les conditions de lyse préalablement décrites doivent donc permettre de déstructurer la membrane bactérienne, mais pas celle des corps d'inclusion. Inversement, quand ces derniers sont isolés, il faut des conditions beaucoup plus drastiques pour solubiliser les agrégats protéiques qu'ils renferment. Le culot issu de la centrifugation du second lavage a été repris dans 5 mL de : Tris-HCl 50 mM, pH 7,5 ; EDTA 10 mM ; β-mercaptoéthanol 6 mM ; Gdn-HCl (puissant agent dénaturant des protéines) 5,6 M, et laissé à 4°C pendant 5 heures. La renaturation a été effectuée par dialyse dans 5 L de : Tris-HCl 50 mM, pH 7,5 ; EDTA 1 mM ; β-mercaptoéthanol 1 mM, puis le dialysat a été centrifugé 10 min à 6000 g. Enfin, le culot de cette dernière centrifugation a été repris dans 5 mL de tampon de dialyse. |
![]()
{kind=link}
{kind=link}